





Food, Supplements, and Smoking: Drug Interaction Insights
Clinical Relevance of Drug Interactions Influenced by Foods, Supplements, and Smoking: A Mechanistic Review
Junichiro Sonoda ¹, Keiki Sakurama ¹, Jin Tokunaga ¹
- First Department of Clinical Pharmacy, School of Pharmaceutical Sciences, Kyushu University of Medical Science, Nobeoka, Miyazaki, Japan
OPEN ACCESS
PUBLISHED: 30 April 2026
CITATION: Sonoda, J., Sakurama, K., et al., 2026. Clinical Relevance of Drug Interactions Influenced by Foods, Supplements, and Smoking: A Mechanistic Review. Medical Research Archives, [online] 14(4).
COPYRIGHT: © 2026 European Society of Medicine. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
ISSN 2375-1924
Abstract
Drug interactions mediated by food, supplements, and smoking are classified as pharmacokinetic interactions (PKDIs) and pharmacodynamic interactions (PDDIs). The interaction between warfarin and natto exemplifies a PDDI, as vitamin K synthesized by fermenting bacteria in natto attenuates anticoagulant efficacy, with a delayed onset and prolonged duration of effect. Grapefruit juice–drug interactions are well characterized and are primarily mediated by irreversible, mechanism-based inhibition of intestinal cytochrome P450 3A4 (CYP3A4) by certain constituents of grapefruit juice. This interaction results in several fold increases in the area under the concentration–time curve (AUC) and maximum plasma concentration (Cmax) of CYP3A4 substrates, such as felodipine. Furthermore, grapefruit juice inhibits intestinal organic anion transporting polypeptides (OATP) 1A2 mediated uptake, thereby decreasing exposure to drugs such as fexofenadine. Similarly, other fruit juices, including orange and apple juices, reduce drug absorption by inhibiting intestinal OATPs using their constituent compounds. Green tea significantly reduces exposure to substrate drugs via competitive inhibition of intestinal OATP1A2 by epigallocatechin gallate (EGCG), a green tea constituent. St John’s wort induces CYP3A4 and P-glycoprotein (P-gp) via activation of the pregnane X receptor by its principal active constituent, substantially reducing plasma concentrations of drugs that are substrates of CYP3A4 and P-gp. Garlic supplements exhibit antiplatelet properties and can cause PDDIs by suppressing platelet aggregation through the modulation of key pathways involved in platelet activation. They are also known to produce PKDIs with HIV-1 protease inhibitors. Cigarette smoking contributes to clinically meaningful interactions, as smokers often exhibit reduced plasma concentrations of drugs that are CYP1A2 substrates, leading to attenuated therapeutic effects and increased toxicity. In oncology, smokers receiving taxanes, gemcitabine, or irinotecan often demonstrate reduced hematologic toxicity, which reflects diminished antitumor efficacy. Collectively, these examples highlight the diverse mechanisms, temporal characteristics, and clinical implications of food, supplement, and smoking-mediated drug interactions.
Keywords
Drug interactions, pharmacokinetics, pharmacodynamics, food, supplements, smoking
Introduction
The number of medicines currently approved for therapeutic use worldwide is estimated to be approximately 4,800. Ensuring the efficacy and safety of these agents is a fundamental responsibility of healthcare professionals. Even when medications are prescribed and administered appropriately, unexpected adverse effects and diminished therapeutic responses may occur. Among the various contributing factors, drug–drug and food–drug interactions represent important and often under-recognized sources of variability in therapeutic outcomes.
Food–drug interactions are frequently insufficiently documented in package inserts, and clinically relevant information is not always readily available. Moreover, even when such interactions occur, changes in drug exposure are rarely verified via therapeutic drug monitoring, raising the possibility that clinically meaningful interactions may be undetected. The potential for encountering previously unrecognized interactions further complicates clinical decision-making.
Grapefruit juice–drug interactions are widely known, and many involve enhanced drug effects due to the inhibition of drug-metabolizing enzymes. Conversely, reduced drug efficacy may result from the inhibition of intestinal drug-transporting proteins, illustrating the diverse mechanisms through which food components can alter pharmacokinetics. Understanding these mechanisms can provide valuable insights into the onset, duration, and magnitude of interaction effects. This, in turn, can improve the ability to anticipate and manage interactions in clinical practice.
Food–drug interactions may occur when constituents present in foods influence metabolic enzymes or transporters, thereby altering the exposure of co-administered medications, or when they modify pharmacological responses through shared or distinct mechanisms of action. However, because some food–drug interactions remain mechanistically unexplained, it is not feasible to provide an exhaustive list of all the potential interactions. In this review, we focus on the interactions between medications and commonly consumed beverages, dietary supplements, and lifestyle-related substances, for which the underlying mechanisms have been characterized. We summarize the available evidence and discuss how healthcare professionals can recognize, evaluate, and interpret these interactions when they encounter them in clinical practice.
Pharmacokinetic Interactions and Pharmacodynamic Interactions
Drug interactions are classified into pharmacokinetic (PKDIs) and pharmacodynamic (PDDIs) interactions. PKDIs occur when a concomitant drug (or dietary component) affects another drug during its absorption, distribution, metabolism, or excretion, thereby altering drug concentrations at the site of action and consequently enhancing or diminishing the therapeutic effects. These interactions include chemical effects on xenobiotic-metabolizing enzymes and transporters that determine the time course of the concentration of one or both chemicals in the body.
In contrast, PDDIs occur when a concomitant drug (or dietary component) with similar pharmacological activity produces additive or synergistic effects or when a concomitant drug (or dietary component) with opposing pharmacological effects reduces the pharmacological response. Predicting pharmacodynamic interactions and their clinical consequences requires identifying the biological sites and mechanisms of action of both substances and determining whether sufficient concentrations are achieved at the sites of action.
When the time to onset, duration, and magnitude of an interaction can be anticipated, the interaction can be avoided, or its impact can be minimized. Pharmacokinetic interactions are relatively straightforward to predict and manage because they are typically accompanied by measurable changes in pharmacokinetic parameters, including the area under the concentration–time curve (AUC) and maximum plasma concentration (Cmax). In contrast, PDDIs arise from diverse mechanisms and causes, making them difficult to predict and challenging to manage.
The principles used to evaluate drug–drug interactions should also be applied when identifying potential supplement-induced interactions with other bioactive compounds, including foods, beverages, and other dietary supplement ingredients.
Natto–Warfarin Interaction: Mechanistic Features of Pharmacodynamic Interactions
Natto is a traditional Japanese fermented soybean product produced using Bacillus subtilis var. natto. In addition, B. subtilis var. natto synthesizes vitamin K. Natto, therefore, contains high concentrations of this vitamin, and bacteria reaching the intestine after ingestion can continue producing vitamin K in vivo, resulting in elevated plasma vitamin K levels that may persist for several days.
The interaction between warfarin and natto exemplifies a PDDI that is difficult to predict. Warfarin exerts its anticoagulant effect by interfering with the cyclic interconversion of vitamin K and its 2,3-epoxide (vitamin K epoxide) through inhibition of vitamin K epoxide reductase, thereby limiting the γ-carboxylation of vitamin K–dependent coagulation proteins. Consequently, the simultaneous consumption of warfarin and natto can substantially attenuate the anticoagulant efficacy of warfarin. Notably, the interaction becomes apparent only after several days, persisted for several days, and is observed exclusively in vivo
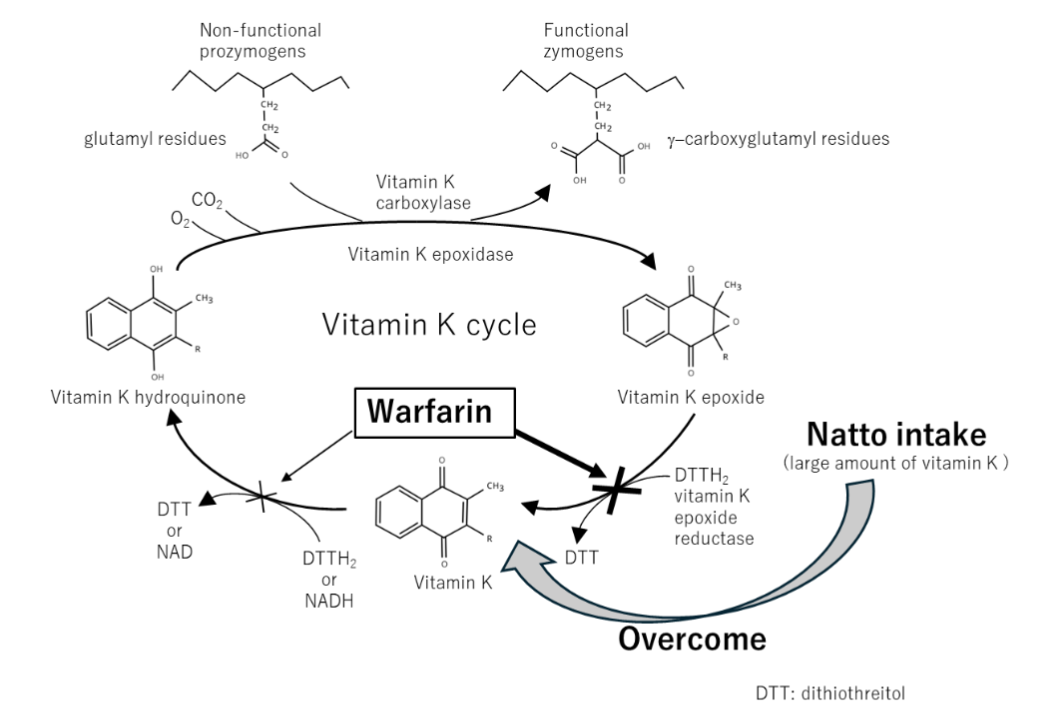
.
Grapefruit Juice ‑Mediated Interactions Involving Cytochrome P450 3A4
Drug‑Metabolizing Enzyme – grapefruit juice–drug interactions: Grapefruit juice–drug interactions involving the metabolic enzyme cytochrome P450 3A4 (CYP3A4) are widely recognized and well-characterized. CYP3A4 is expressed in the apical enterocytes of the small intestine and hepatocytes in the liver. Cytochrome P450 enzymes, particularly CYP3A4, participate in the metabolism of approximately half of all clinically used drugs, including calcium channel blockers, statins, immune-suppressants, benzodiazepines, and antihistamines. These interactions were initially identified in an interventional study by Bailey et al., which investigated the hemodynamic interaction between ethanol and felodipine. White grapefruit juice was incidentally used to mask the taste of ethanol, leading to the unexpected identification of a potential interaction. A decade later, Bailey et al. published a comprehensive review describing grapefruit juice–drug interactions.
Co-administration of felodipine with grapefruit juice showed a pronounced interaction, increasing Cmax by several-fold compared with administration with water, enhancing the hemodynamic effects of felodipine and resulting in clinically relevant adverse events. A total of 5 days of grapefruit juice consumption did not alter hepatic CYP3A4 activity but reduced CYP3A4 protein content in small-bowel enterocytes by a mean of 62%, which was associated with more than 3-fold and 5-fold increases in the AUC and Cmax of felodipine, respectively. Therefore, grapefruit juice exerts minimal influence on hepatic CYP3A4 activity, and its pharmacokinetic effects are generally limited to orally administered drugs. Consequently, the interaction is considered negligible for administration routes that bypass intestinal absorption, including intravenous dosing. However, ingestion of large quantities of grapefruit juice (double-strength grapefruit juice, 240 mL thrice daily for a total of 9 administrations) inhibited CYP3A4 not only in the intestine but also in the liver, resulting in prolongation of the elimination half-life. These findings suggest that high grapefruit juice intake may modulate drug disposition even when drugs are administered via non-oral routes.
The magnitude of grapefruit–drug interactions varies among agents of the same pharmacological class. For example, among dihydropyridine calcium channel blockers, co-administration with grapefruit juice increased drug Cmax by a mean of 406% (range, 107–836%) for nisoldipine, whereas the increase for amlodipine was 115% (79–165%) relative to administration with water. This difference is explained by the fact that dihydropyridine agents with low intrinsic oral bioavailability exhibit greater interaction magnitude with grapefruit juice than agents with higher oral bioavailability. The oral bioavailability of amlodipine and nisoldipine is approximately 60 and 5%, respectively. Amlodipine undergoes less first-pass metabolism than other dihydropyridines, which partly explains its more stable pharmacokinetic profile. In addition, amlodipine exhibits high tissue affinity and undergoes slow absorption and distribution, resulting in a time to maximum concentration of 6–12 h, and its pharmacokinetics are largely unaffected by food intake. In contrast, the CYP3A4-inhibitory constituents of grapefruit juice undergo glucuronidation after ingestion, with a reported median onset of glucuronide excretion approximately 2 h after the administration of these compounds. These pharmacokinetic differences likely contribute to the minimal effect of grapefruit juice on amlodipine.
Notably, grapefruit juice–drug interactions involving drugs with low oral bioavailability exhibit substantial inter-individual variability. Some patients experience pronounced effects, whereas others show minimal or no responses. CYP3A4 content in the small intestine varies by at least 10-fold among individuals, and because grapefruit juice–drug interactions involve reductions in CYP3A4 protein levels, this variability directly influences the magnitude of the interaction. Consistent with this mechanism, reductions in enterocyte CYP3A4 concentrations after 5 days of grapefruit juice ingestion correlate with increases in Cmax of felodipine.
Temporal characteristics of the grapefruit juice–drug interactions: Several studies have described the temporal onset and duration of grapefruit juice–drug interactions. Lundahl et al. investigated increases in the Cmax and AUC of felodipine when 200 mL of grapefruit juice was administered at different intervals before felodipine dosing. When felodipine was administered concurrently with grapefruit juice (0 h interval) or after a 4 h interval, the AUC increased by approximately 45% compared to that in the control group. As the dosing interval increased, the interaction gradually diminished, resulting in a 25% increase at a 10 h interval. Even at a 24 h interval, Cmax remained significantly elevated by 32%. These findings demonstrate that the interaction occurs immediately after co-administration and that the effect of grapefruit juice persists for at least 24 h. In another study, the consumption of one glass (8 oz) of grapefruit juice increased the AUC and Cmax of the drug by 225 and 216%, respectively, compared with the control. When the same amount of grapefruit juice was consumed three times daily for five consecutive days, the AUC and Cmax increased by an additional 34 and 44%, respectively, beyond the values observed after a single ingestion. Grapefruit juice intake is accompanied by a reduction in enterocyte CYP3A4 protein levels, indicating that grapefruit juice acts as an irreversible mechanism-based inhibitor of CYP3A in humans.
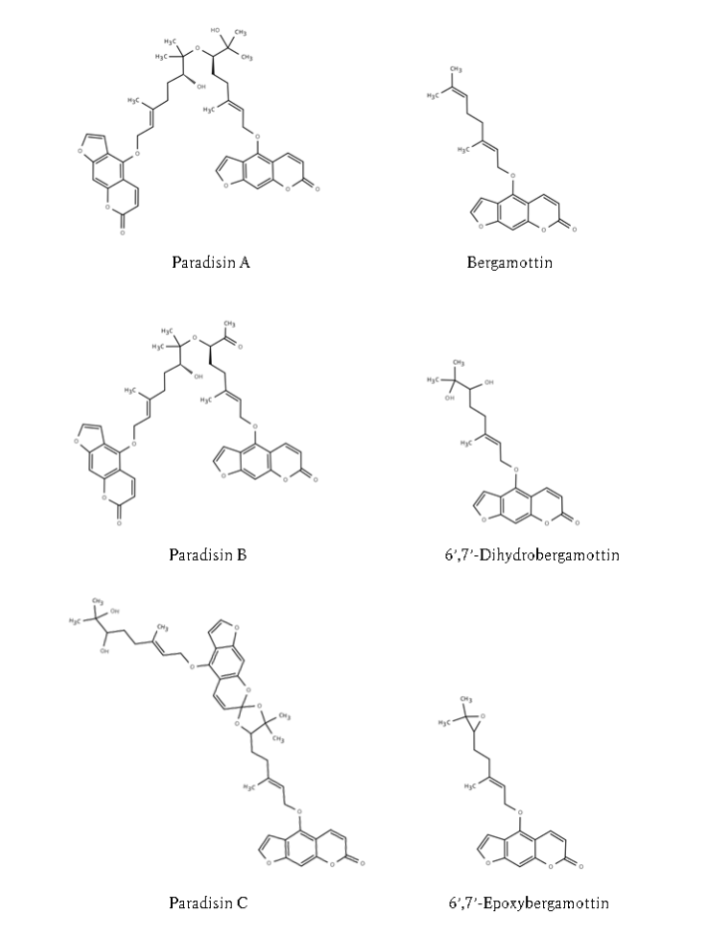
Constituents of grapefruit responsible for drug interactions: Furanocoumarins and flavonoids are the key constituents responsible for grapefruit juice–drug interactions. The furanocoumarins present in grapefruit include bergamottin, 6’,7’-dihydroxybergamottin, 6’,7’-epoxybergamottin, and the dimers parasihin A, parasihin B, and parasihin C. Flavonoid constituents include glycosides such as naringin and hesperidin. Two independent investigations examined the active constituents by separating grapefruit juice samples into precipitate and supernatant fractions by centrifugation and comparing their respective components. The precipitate fraction contained bergamottin, 6’,7’-epoxybergamottin, paradisin A, and paradisin B, whereas the supernatant fraction predominantly contained 6’,7’-dihydroxybergamottin and naringin. When constituents from each fraction were evaluated for their capacity to increase AUC and Cmax in healthy volunteers or to inhibit CYP3A4 activity, the precipitate fraction consistently exerted stronger effects than the supernatant. Although naringin is an abundant constituent of grapefruit juice, it is unlikely to contribute substantially to CYP3A4 inhibition. Among the furanocoumarins, the dimeric species are markedly stronger inhibitors of CYP3A4 activity than their monomeric counterparts. The inhibitory potency of grapefruit juice samples also tends to increase with higher total furanocoumarin content.
Other citrus fruits besides grapefruit, including sweetie (Oroblanco), pomelo, sour orange, and Seville orange, also contain multiple furanocoumarins that may act synergistically to produce interactions with CYP3A4 substrate drugs. In addition, Banpeiyu, Hassaku orange, and Kumquat, which are commonly consumed in Japan, exhibit inhibitory activity toward CYP3A4 in vitro.
Grapefruit Juice and Other Beverage‑Mediated Interactions Involving Organic Anion-Transporting Polypeptide Transports
Grapefruit juice–fexofenadine: As discussed above, grapefruit juice acts as an irreversible mechanism-based inhibitor of human CYP3A, thereby increasing the bioavailability of CYP3A substrate drugs. In contrast, grapefruit juice intake decreased the AUC of fexofenadine by approximately 55% compared to that of water. They also reported the effects of the particle fraction obtained from centrifuged grapefruit juice, which increases the AUC of felodipine through CYP3A4 inhibition and naringin, a major constituent of grapefruit juice. The AUC values for the particle fraction and naringin were 97 and 75%, respectively. Furthermore, in a [³H]-fexofenadine uptake assay using HeLa cells transiently expressing the uptake transporter organic anion-transporting polypeptide (OATP) 1A2, naringin inhibited uptake to a degree similar to that of verapamil, a positive control for OATP1A2 inhibition (IC₅₀: verapamil, 2.6 μM; naringin, 3.6 μM). These findings indicate that the interaction between grapefruit juice and fexofenadine results from the inhibition of OATP1A2-mediated fexofenadine uptake by naringin present in grapefruit juice.
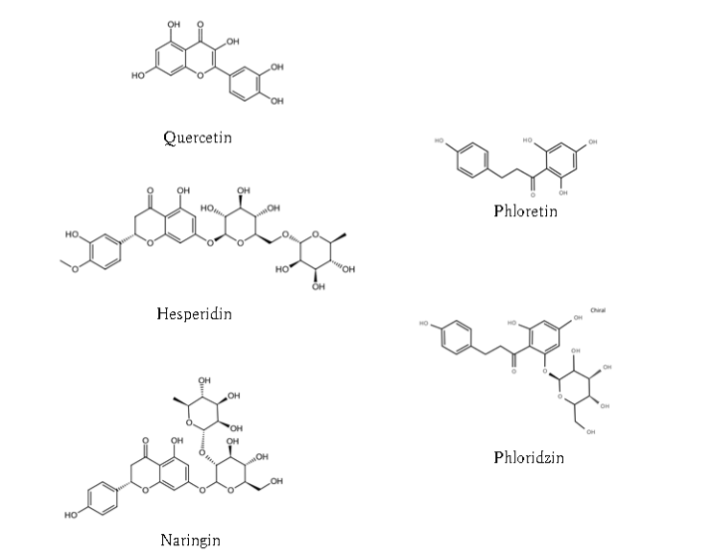
Fruit juices (other than grapefruit juice)–fexofenadine: Dresser et al. examined interactions between fexofenadine and other fruit juices, including orange and apple juices. Similar to grapefruit juice, co-administration of orange or apple juice decreased the AUC, urinary excretion, and Cmax of fexofenadine by approximately 20–40% relative to water. Consistent with these findings, [14C]‑fexofenadine uptake in OATP-expressing cells was markedly reduced by 20-fold-diluted grapefruit, orange, and apple juices. The major bioactive constituents of fruit juices include naringin, hesperetin, phloridzin (a glucoside), phloretin (an aglycone), hesperidin, and quercetin. Naringin, hesperidin, and quercetin are flavonoids, whereas phloridzin and phloretin are dihydrochalcones.
Green tea−nadolol: Misaka S. and colleagues reported an OATP1A2-mediated interaction between green tea and nadolol, a β-adrenergic receptor blocker. After 14 days of green tea consumption (700 mL/day), the administration of a single 30-mg nadolol tablet produced marked reductions in nadolol exposure, with AUC and Cmax decreasing to 14.7 and 15.0%, respectively, relative to water intake. In vitro [³H]-nadolol uptake assays using human embryonic kidney 293 (HEK293) cells stably expressing OATP1A2 or OATP2B1, nadolol was identified as a substrate of OATP1A2 but not OATP2B1. Furthermore, epigallocatechin gallate (EGCG), a major flavan-3-ol constituent of green tea, inhibited OATP1A2-mediated [³H]-nadolol uptake in a concentration-dependent manner, with an IC₅₀ of 37.3 μM. Because the concentration of EGCG in green tea is typically approximately 1,000 μM, EGCG present in the beverage could inhibit OATP1A2-mediated nadolol uptake in enterocytes even after up to a 27-fold dilution by gastrointestinal fluids.
Epigallocatechin gallate-mediated inhibition of the transport function of intestinal organic anion-transporting polypeptides: Abe et al. conducted inhibition kinetic analyses using Dixon plots with multiple concentrations of nadolol as a substrate for OATP1A2 to characterize the inhibitory mechanism of EGCG. EGCG competitively inhibited OATP1A2-mediated uptake of nadolol with a Ki value of 19.4 μM. Hence, EGCG may also act as a substrate for OATP1A2. Consistent with this interpretation, EGCG is transported by OATP1A2 with a Km value of 18.8 μM. In contrast, Misaka et al. reported a Km value of 84.3 μM for OATP1A2-mediated transport of nadolol, which is more than four-fold higher than that of EGCG. Thus, EGCG exhibits a substantially higher affinity for OATP1A2 than nadolol. This higher affinity, together with the competitive inhibition of OATP1A2, is consistent with the marked reduction in nadolol plasma concentrations to approximately 15% when green tea is co-administered, as described above.
Because OATP1A2 and OATP2B1 are localized on the apical membrane of enterocytes, interactions among orally administered substrates are likely to occur at high local concentrations at these transporter sites, producing direct and substantial effects. Epigallocatechin gallate inhibits both OATP1A2 and OATP2B1 in vitro. Accordingly, drugs that are substrates of both OATP1A2 and OATP2B1, such as fexofenadine, celiprolol, and aliskiren, are expected to exhibit pronounced interactions when co-administered with green tea. Evidence supporting this prediction has been reported for celiprolol. We previously demonstrated that green tea consumption markedly reduced the AUC and urinary excretion of celiprolol by 98.6 and 98.0%, respectively, compared with water. A plausible explanation for the near elimination of celiprolol exposure is that green tea components, most likely EGCG, act as substrates for intestinal OATP1A2 and OATP2B1 with a higher affinity than celiprolol. EGCG may therefore be preferentially transported by these intestinal OATPs, resulting in strong inhibition of celiprolol uptake and absorption.
Temporal characteristics of the intestinal organic anion-transporting polypeptide-mediated interactions: Intestinal OATP-mediated interactions occur at the apical membrane of enterocytes in the small intestine, where OATP1A2 and OATP2B1 are expressed. When a substrate drug for these transporters and another compound with a higher transporter affinity are present simultaneously at this site, the drug substrate is likely to undergo competitive inhibition, resulting in reduced uptake (absorption).
Dolton et al. clearly described the characteristics of CYP3A4-mediated interactions induced by grapefruit juice and those of intestinal OATP-mediated interactions caused by fruit juices. Interactions between grapefruit juice and CYP3A4 substrates occur through an irreversible mechanism in which CYP3A4 protein in small intestinal enterocytes is reduced without corresponding changes in CYP3A4 mRNA expression. For some drugs, the magnitude of this interaction decreases gradually over several days. In contrast, interactions mediated by grapefruit juice through intestinal OATPs do not result from reductions in transporter protein levels but from the inhibition of transporter-mediated uptake. Glaeser et al. reported that duodenal biopsy samples obtained 1–2 h after grapefruit juice ingestion showed no significant differences in OATP1A2 protein or SLCO1A2 mRNA levels compared with those observed after water ingestion. However, systemic exposure to fexofenadine, an OATP1A2 substrate, decreased by approximately half when co-administered with grapefruit juice compared to that with water. Notably, the inhibition of OATPs by grapefruit juice resolved more rapidly than the inhibition of CYP3A4 caused by grapefruit juice. These findings indicate that grapefruit juice–mediated OATP interactions may be avoided by separating medication and juice intake by at least 4 h.
Interactions mediated by intestinal OATPs (OATP1A2/2B1) can have direct and substantial effects because they occur at the apical membrane of small intestinal enterocytes, where locally high concentrations of co-administered compounds are achieved. However, the current drug interaction guidelines issued by major regulatory authorities—including the Food and Drug Administration (FDA), European Medicines Agency (EMA), and Pharmaceuticals and Medical Devices Agency (PMDA)—primarily address hepatic uptake transporters, such as OATP1B1 and OATP1B3, whereas intestinal transporters, such as OATP1A2 and OATP2B1, are not specifically addressed. Consequently, clinical awareness of intestinal OATP-mediated interactions remains limited in current medical practice.
St John’s Wort
St John’s Wort−drug interactions: St John’s wort is widely used as an herbal medicine for the management of mild to moderate depressive episodes. In a systematic review, Linde et al. summarized the evidence from placebo-controlled trials and comparisons with conventional antidepressants. Among placebo-controlled studies, notable differences were observed between large and small trials: in nine large trials, the response rate ratio (RR) for St John’s wort extracts compared with placebo was 1.28 (95% CI, 1.10–1.49), whereas in nine smaller trials, the RR was 1.87 (95% CI, 1.22–2.87). In contrast, trials comparing St John’s wort extract with conventional antidepressants showed statistical homogeneity. Compared with tricyclic or tetracyclic antidepressants and selective serotonin reuptake inhibitors (SSRIs), the corresponding RRs were 1.02 (95% CI, 0.90–1.15; five trials) and 1.00 (95% CI, 0.90–1.11; 12 trials), respectively. Notably, treatment discontinuation due to adverse effects occurred less frequently in patients receiving St John’s wort extract than in those receiving conventional antidepressants.
While St John’s wort is marketed as a dietary supplement in many countries, standardized dry extract formulations are used as phytomedicine-regulated medicinal products to treat mild-to-moderate depressive episodes. However, the principal concern with St John’s wort is the reduction in the therapeutic efficacy of concomitant medications caused by drug interactions. In 2000, the European Medicines Evaluation Agency (EMEA) issued a Public Statement warning of interactions between St John’s wort products and indinavir, noting that subtherapeutic plasma concentrations of indinavir may lead to antiretroviral resistance and treatment failure, with serious clinical consequences for patients with HIV-1 infection.
Clinical reports and insights on St John’s Wort–drug interactions: As highlighted in the aforementioned Public Statement, these interactions are largely attributed to the induction of the cytochrome P450 isoenzyme CYP3A4 by St John’s wort. St John’s wort has also been shown to induce the efflux transporter P-glycoprotein (P-gp). Consequently, drugs that are substrates of CYP3A4 and/or P-gp are susceptible to interactions with St John’s wort, leading to decreased systemic exposure.
In a clinical intervention study involving renal transplant recipients receiving tacrolimus or cyclosporine A, the administration of St John’s wort extract for 14 days resulted in marked reductions in drug exposure. For tacrolimus, the dose required to maintain therapeutic concentrations increased from 4.5 to 8.0 mg/day, corresponding to a 58% reduction in the dose-corrected AUC. For cyclosporine A, increasing the dose from 2.7 to 4.2 mg/kg/day was accompanied by a 41% reduction in dose-corrected AUC.
The study also evaluated the pharmacokinetic changes in five cyclosporine A metabolites (AM1, AM1C, AM9, AM19, and AM4N) after St John’s wort administration. The metabolites displayed two distinct patterns of distribution. In the uncorrected data, the AUCs of AM1 and AM1C decreased by 35–40%, whereas those of AM9, AM19, and AM4N increased by 20–51%. Cyclosporine A and its metabolites are primarily eliminated via biliary excretion, a process strongly influenced by P-gp–mediated transport. These differences likely reflect variations in metabolite affinity for P-gp. Metabolites with a high affinity for P-gp are expected to undergo enhanced biliary excretion and reduced enterohepatic recirculation, resulting in lower plasma concentrations (AM1 and AM1C). In contrast, metabolites that are weaker P-gp substrates are more strongly influenced by CYP3A4 induction by St John’s wort, leading to increased plasma concentrations (AM9, AM19, and AM4N).
Mechanisms and extent of St John’s Wort–mediated drug interactions: Moore et al. investigated the pregnane X receptor (PXR), a principal regulator of CYP3A4 transcription belonging to the steroid/thyroid hormone nuclear receptor family, using human hepatocytes transfected with PXR and provided clear evidence for the mechanism by which St John’s wort induces CYP3A4. Hyperforin, the principal active constituent of St John’s wort, binds to PXR in competitive ligand-binding assays and exhibits potent PXR activation (EC₅₀ = 23 nM), exceeding that of the established PXR agonist rifampicin. Through this strong PXR activation, hyperforin induces CYP3A4 transcription. According to Biber et al., a single dose of standardized St John’s wort extract (300 mg) produced a peak plasma hyperforin concentration of 285 nM, whereas steady-state concentrations reached approximately 480 nM after repeated administration of 300 mg three times daily for eight days. On the basis of these findings, the authors concluded that St John’s wort activates PXR in humans.
The magnitude of CYP3A4 induction associated with St John’s wort can be contextualized by comparing it with classical CYP3A4 inducers, such as rifampin and carbamazepine. St John’s wort reduced the elimination half-life of alprazolam from 12.4 to 6.0 h. By comparison, carbamazepine reduced the half-life from 17.1 to 7.7 h, whereas rifampin produced a more pronounced reduction from 14.1 to 2.6 h. St John’s wort induces the expression of both CYP3A4 and P-gp. In addition, PXR activation has been shown to induce P-gp expression. Because P-gp and CYP3A4 exhibit distinct but overlapping substrate profiles, the magnitude of clinically relevant interactions depends on whether a drug is predominantly transported by P-gp, metabolized by CYP3A4, or subject to both pathways. Tacrolimus and cyclosporine A are substrates of both CYP3A4 and P-gp, and the observed interactions are therefore presumed to involve contributions from both mechanisms. However, the relative contribution of each pathway may vary depending on the drug involved, inter-individual variability, and underlying disease states.
Garlic Supplements
Garlic supplements and interactions with conventional medications: Garlic, a nutraceutical herb rich in organosulfur compounds, has been consumed since ancient times for its health-promoting properties. These organosulfur compounds are responsible for the characteristic taste and odor of garlic and play a central role in mediating its pharmacological effects. Garlic has demonstrated preventive and therapeutic potential against a broad spectrum of non-communicable diseases—including cancer, cardiovascular diseases, metabolic disorders, hypertension, and diabetes—through mechanisms involving antioxidant, anti-inflammatory, and lipid-lowering activities, as demonstrated in numerous in vitro, in vivo, and clinical studies.
Garlic supplement–mediated pharmacodynamic drug interactions: Among the various health benefits of garlic, particular attention has been directed toward its effects on cardiovascular diseases. Current evidence indicates that garlic exerts beneficial effects on atherosclerosis, hypertension, and diabetes, thereby contributing to a reduced risk of myocardial infarction and ischemic stroke. Garlic supplements possess antiplatelet activity that may contribute to attenuation of cardiovascular risk factors. However, pharmacodynamic drug interactions (PDDIs) may be noted depending on concomitant medications.
Using a platelet-rich plasma (PRP) assay, Steiner et al. demonstrated that six weeks of aged garlic extract supplementation dose-dependently suppressed platelet aggregation induced by epinephrine, adenosine diphosphate (ADP), and collagen, as well as platelet adhesion to collagen, fibrinogen, and von Willebrand factor. Similarly, Rahman et al. reported that platelets obtained after 13 weeks of aged garlic extract supplementation exhibited significant reductions in both the total percentage and initial rate of ADP-induced platelet aggregation. The Km value for ADP-induced aggregation increased approximately two-fold after supplementation.
Two mechanisms have been proposed to explain the inhibitory effects of aged garlic extract on platelet aggregation. The first mechanism involves the inhibition of fibrinogen binding to the platelet membrane glycoprotein GPIIb/IIIa complex. During ADP-mediated platelet activation in the presence of extracellular Ca²⁺, the GPIIb/IIIa heterodimer binds to fibrinogen to form cross-links required for platelet aggregation. This interaction is inhibited by ajoene, a garlic-derived organosulfur compound, owing to the abundance of sulfhydryl groups in the GPIIb/IIIa receptor. The second proposed mechanism involves the modulation of intracellular calcium signaling. Activation of platelet ADP receptors normally increases cytosolic Ca²⁺ levels, promoting platelet aggregation. Because aged garlic extract supplementation inhibited ADP-induced aggregation in this study, the extract may also interfere with the ADP-mediated increase in intracellular Ca²⁺. Indeed, garlic has been suggested to suppress intracellular Ca²⁺ mobilization in platelets either by inhibiting Ca²⁺ influx through chelation within the platelet cytosol or by modulating other intracellular second-messenger pathways. Therefore, garlic supplementation may contribute to a reduction in cardiovascular risk factors. However, in patients receiving antiplatelet therapy, prolonged garlic supplementation may potentiate antiplatelet activity and increase the risk of bleeding.
A review of the literature indicates that garlic intake predisposes patients to pathological bleeding, and a recent systematic review provided strong evidence that garlic consumption increases the risk of surgical bleeding. Although concomitant use of garlic with warfarin did not increase prothrombin time–international normalized ratio (PT–INR) values, garlic significantly reduced platelet aggregation, a pharmacodynamic effect that may directly increase the bleeding risk. Numerous in vitro studies and case reports also indicate that garlic can directly promote bleeding. Consequently, PDDIs between garlic supplements and antiplatelet agents such as aspirin may markedly potentiate antiplatelet effects and increase the risk of clinically significant bleeding.
Garlic supplement–mediated pharmacokinetic interactions: Garlic supplements have also been reported to affect drug metabolism and transport, particularly in the case of HIV-1 protease inhibitors. In a study involving ten healthy volunteers, the administration of garlic extract capsules (600 mg twice daily; 1,200 mg/day) for 21 days reduced the AUC of saquinavir by 51%. In addition, the maximum (Cmax) and trough (Ctrough) plasma concentrations decreased by 54 and 49%, respectively. Notably, even 10 days after discontinuation of garlic supplementation, the AUC and Cmax of saquinavir recovered only to 60–70% of their baseline levels, suggesting a sustained effect. The authors proposed that this interaction may involve the induction of CYP3A4, resulting in the enhanced metabolism of saquinavir. However, because saquinavir is also a known substrate of P-gp, a contribution from P-gp induction cannot be excluded.
Hajda et al. reported that 21 days of garlic extract administration in healthy volunteers did not alter intestinal CYP3A4 protein levels or hepatic CYP3A4 activity, but increased intestinal P-gp expression by approximately 30%. In addition, although the bioavailability of saquinavir decreased after garlic extract administration, the bioavailability of simvastatin, a known CYP3A4 substrate, remained unchanged. These findings suggest that the altered pharmacokinetics of saquinavir may primarily result from upregulation of intestinal P-gp rather than induction of CYP3A4. In contrast to the pronounced interaction observed with saquinavir, a study in healthy volunteers evaluated the pharmacokinetics of ritonavir after a single dose was administered following four days of garlic extract intake. The AUC of ritonavir decreased by 17%, although this reduction was not statistically significant. Ritonavir undergoes CYP3A4 autoinduction during the first two weeks of therapy; therefore, any induction of P-gp or CYP3A4 caused by garlic intake may have been offset by this autoinduction. These findings suggest that multiple overlapping mechanisms may contribute to the interaction and therefore require cautious interpretation.
Taken together, these observations indicate that the interaction profile of ritonavir during short-term garlic exposure is difficult to interpret because of the presence of several counteracting mechanisms. Therefore, studies conducted under steady-state ritonavir conditions and longer garlic exposure periods are required to clarify the potential influence of garlic on ritonavir pharmacokinetics. Although garlic supplements have been suggested to induce CYP3A4 and/or P-gp activity, the clinical evidence for pharmacokinetic drug interactions involving medications other than HIV-1 protease inhibitors remains limited.
Cigarette Smoking
Current status of tobacco use and associated health impact: Cigarette smoke adversely affects human health. Tobacco use remains one of the most significant global public health threats. The World Health Organization (WHO) characterizes tobacco-related deaths as those attributable to active smoking, secondhand smoke exposure, and the use of tobacco products. According to WHO estimates, more than eight million deaths occur annually worldwide. Although substantial regional variation exists, deaths attributable to smoking were estimated to account for approximately 9.7% of all global deaths in 2023. Tobacco use is more prevalent in men than in women. According to a WHO report (Status of Tobacco Use in the Region), the prevalence of smoking in the WHO European Region in 2025 is estimated to be 30.6% among men and 18.2% among women. Smoking-attributable mortality also shows marked sex differences; among men, smoking accounts for 27% of deaths from non-communicable diseases (NCDs), 25% of deaths from cardiovascular diseases, and 37% of cancer deaths, whereas the corresponding proportions among women are 10, 8, and 15%, respectively. These figures indicate that reducing the smoking prevalence would directly contribute to lowering mortality from major disease categories. Furthermore, tobacco control is positioned as a key strategy for achieving the global target—endorsed at the 2013 World Health Assembly—of “reducing premature mortality from NCDs by 25% by the year 2025.”
Clinical evidence and mechanistic insights into drug interactions caused by cigarette smoking: Cigarette smoking remains a major public health concern because of its extensive adverse effects. Accumulating evidence indicates that smoking alters the efficacy and pharmacokinetics of conventional medications. Therefore, this section examines the mechanistic basis of smoking-related drug interactions and their clinical implications. Cigarette smoke contains numerous harmful chemical constituents, including acetone, benzene, 1,3-butadiene, carbon monoxide, heavy metals, nicotine, and pyridine, as well as polycyclic aromatic hydrocarbons (PAHs), such as benzo[α]pyrene, a compound with well-established carcinogenic properties. PAH metabolism involves enzymes of the CYP1A subfamily, and multiple studies have demonstrated that exposure to PAHs or cigarette smoke induces CYP1A2 expression.
Theophylline, a drug widely used in the management of chronic respiratory diseases, such as bronchial asthma and chronic obstructive pulmonary disease, is primarily metabolized by CYP1A2. Hunt et al. compared the pharmacokinetics of a single oral dose of theophylline between heavy smokers who had smoked 1–2 packs per day for more than one year and nonsmokers with no smoking history for at least two years. The elimination half-life, apparent volume of distribution, and total body clearance in heavy smokers (n = 8) were 4.31 h, 0.50 L/kg, and 100.4 mL/min/1.73 m², respectively, whereas the corresponding values in nonsmokers (n = 8) were 7.03 h, 0.38 L/kg, and 44.5 mL/min/1.73 m², respectively. All pharmacokinetic parameters differed significantly between the two groups. Among the eight heavy smokers, three abstained from smoking for at least three months; however, no appreciable changes in pharmacokinetic parameters were observed. Conversely, three of the eight nonsmokers had a remote smoking history (2, 5, and 6 years previously); however, their pharmacokinetic profiles were indistinguishable from those of the other nonsmokers. These findings suggest that normalization of smoking-related alterations in theophylline pharmacokinetics may require a prolonged period ranging from several months to as long as two years.
The time course over which drug-metabolizing enzyme activity returns to baseline after smoking cessation remains unclear. Powell et al. reported that the mean theophylline elimination half-life in smokers who had abstained for two years was intermediate between that in nonsmokers and current smokers, suggesting that normalization of drug metabolism may require more than two years. In contrast, Piafsky et al. demonstrated that light smokers who had abstained for at least three weeks exhibited a theophylline half-life comparable to that of nonsmokers. In Similarly, a study by Lee et al., in which participants were hospitalized and dietary intake, environmental exposure, and smoking behavior were strictly controlled, showed that theophylline clearance decreased significantly after only one week of smoking cessation.
In addition to theophylline, cigarette smoking has been reported to influence the pharmacokinetics of numerous medications. Zanni et al. conducted a comprehensive review examining the effects of smoking on drug pharmacokinetics and provided important insights into this field. Their analysis showed that, compared with nonsmokers, smokers exhibited significantly lower plasma concentrations of several drugs, including the antipsychotics olanzapine, clozapine, quetiapine, and risperidone; the antidepressants escitalopram, amitriptyline, doxepin, and mirtazapine; the antihypertensive agent verapamil; the antibiotic metronidazole; and the anticancer agents irinotecan and erlotinib.
Smoking-related alterations in drug exposure may also affect the therapeutic outcomes. For example, improvement in psychiatric symptoms during olanzapine therapy has been reported to be less pronounced in smokers than in nonsmokers. In addition, hematological toxicity associated with anticancer agents such as taxanes (docetaxel and paclitaxel), gemcitabine, and irinotecan has been reported to be less severe in smokers than in nonsmokers. Reduced hematological toxicity in smokers may reflect lower systemic drug exposure, which could potentially compromise antitumor efficacy and increase the risk of treatment failure.
Conclusion
Understanding the diverse mechanisms through which foods, beverages, dietary supplements, and lifestyle factors influence drug disposition and therapeutic responses is essential for ensuring safe and effective pharmacotherapy. Classic examples, such as the attenuation of the anticoagulant effect of warfarin by vitamin K–rich natto, illustrate the potential for profound pharmacodynamic consequences. Grapefruit juice and other fruit juices demonstrate how the modulation of intestinal CYP3A4 or OATP transporters can markedly alter systemic drug exposure. St John’s wort exemplifies how herbal supplements can reduce therapeutic drug concentrations through induction of CYP3A4 and P-gp. Garlic supplements add further complexity by exerting antiplatelet effects and potentially influencing CYP3A4 and/or P-gp expression, thereby posing both pharmacodynamic and pharmacokinetic risks. Cigarette smoking is an additional lifestyle factor that reduces the plasma concentrations of CYP1A2 substrate drugs and may attenuate both therapeutic effects and drug-related toxicities, with implications for psychiatric and oncological treatments.
Future perspectives include advancing the systematic evaluation of food and supplement constituents using standardized experimental models, establishing frameworks that incorporate transporter-mediated mechanisms into interaction assessments, and developing predictive tools that account for inter-individual variability. Enhancing pharmacovigilance efforts and accumulating real-world evidence are essential for identifying interactions that have not yet been recognized. Ultimately, enhancing both mechanistic understanding and clinical awareness of these interactions will contribute to safer and more effective pharmacotherapy, enabling healthcare professionals to anticipate risks, optimize drug therapy, and provide evidence-based guidance to patients living in increasingly diverse dietary and lifestyle environments.
Conflicts of Interest Statement
The authors confirm that the content of this article has no conflicts of interest.
Funding Statement
We gratefully acknowledge the funding from the Kyushu University of Medical Science.
Acknowledgments
We thank Ms. Tsuchimochi for assistance with literature organization. We also thank Editage for editing and reviewing this manuscript for English language.
References
- Knox C, Wilson M, Klinger CM, et al. DrugBank 6.0: the DrugBank Knowledgebase for 2024. Nucleic Acids Res. 2024;52(D1):D1265–D1275.
- National Academies of Sciences, Engineering, and Medicine. Dietary Supplements: A Framework for Evaluating Safety. Washington, DC: The National Academies Press; 2005: 236-239.
- Homma K, Wakana N, Suzuki Y, et al. J Nutr Sci Vitaminol (Tokyo), 2006;52(5):297-301.
- Hirsh J, Fuster V, Ansell J, Halperin JL. American Heart Association; American College of Cardiology Foundation. Circulation. 2003;107(12):1692-1711.
- Aosaki M, Iwade K, Echizen H, eds. Information on rational use of Warfarin. 3rd ed. Tokyo: Eisai Co., Ltd.; 2006:18.
- Aosaki M. Oral anticoagulants. J Cardiol Dept. 1981;10(3):218–228.
- Nutescu EA, Burnett A, Fanikos J, Spinler S, Wittkowsky A. Erratum to: Pharmacology of anticoagulants used in the treatment of venous thromboembolism. J Thromb Thrombolysis. 2016;42(2):296-311.
- Moore LB, Goodwin B, Jones SA, el al. St. John’s wort induces hepatic drug metabolism through activation of the pregnane X receptor. Proc Natl Acad Sci U S A. 2000;97(13):7500-7502.
- Dayyih WA, Al-Ani I, Hailat M, et. al. Review of grapefruit juice-drugs interactions mediated by intestinal CYP3A4 inhibition. J. Appl. Pharm. Sci. 2024;14(5):59-68.
- Bailey DG, Spence JD, Edgar B, Bayliff CD, Arnold JM. Ethanol enhances the hemodynamic effects of felodipine. Clin Invest Med. 1989;12(6):357-362.
- Bailey DG, Malcolm J, Arnold O, Spence JD. Grapefruit juice-drug interactions. Br J Clin Pharmacol. 1998;46(2):101-110.
- Blychert E, Edgar B, Elmfeldt D, Hedner T. A population study of the pharmacokinetics of felodipine. Br J Clin Pharmacol. 1991;31(1):15-24.
- Bailey DG, Bend JR, Arnold JM, Tran LT, Spence JD. Erythromycin-felodipine interaction: magnitude, mechanism, and comparison with grapefruit juice. Clin Pharmacol Ther. 1996;60(1):25-33.
- Bailey DG, Arnold JM, Munoz C, Spence JD. Grapefruit juice–felodipine interaction: reproducibility and characterization with the extended release drug formulation. Br J Clin Pharmacol. 1995;40(2):135-140.
- Edgar B, Bailey D, Bergstrand R, Johnsson G, Regårdh CG. Acute effects of drinking grapefruit juice on the pharmacokinetics and dynamics of felodipine–and its potential clinical relevance. Eur J Clin Pharmacol. 1992;42(3):313-317.
- Bailey DG, Arnold JM, Munoz C, Spence JD. Grapefruit juice–felodipine interaction: mechanism, predictability, and effect of naringin. Clin Pharmacol Ther. 1993;53(6):637-642.
- Lown KS, Bailey DG, Fontana RJ, et. al. Grapefruit juice increases felodipine oral availability in humans by decreasing intestinal CYP3A protein expression. J Clin Invest. 1997;99(10):2545-2553.
- Ducharme MP, Warbasse LH, Edwards DJ. Disposition of intravenous and oral cyclosporine after administration with grapefruit juice. Clin Pharmacol Ther. 1995;57(5):485-491.
- Veronese ML, Gillen LP, Burke JP, et. al. Exposure-dependent inhibition of intestinal and hepatic CYP3A4 in vivo by grapefruit juice. J Clin Pharmacol. 2003;43(8):831-839.
- Bailey DG, Arnold JM, Strong HA, Munoz C, Spence JD. Effect of grapefruit juice and naringin on nisoldipine pharmacokinetics. Clin Pharmacol Ther. 1993;54(6):589-594.
- Josefsson M, Zackrisson AL, Ahlner J. Effect of grapefruit juice on the pharmacokinetics of amlodipine in healthy volunteers. Eur J Clin Pharmacol. 1996;51(2):189-193.
- Stopher DA, Beresford AP, Macrae PV, Humphrey MJ. The metabolism and pharmacokinetics of amlodipine in humans and animals. J Cardiovasc Pharmacol. 1988;12(sup7):S55-59.
- Dudhipala N, Janga KY, Gorre T. Comparative study of nisoldipine-loaded nanostructured lipid carriers and solid lipid nanoparticles for oral delivery: preparation, characterization, permeation and pharmacokinetic evaluation. Artif Cells Nanomed Biotechnol. 2018;46(sup2):616-625.
- Lown KS, Kolars JC, Thummel KE, et al. Interpatient heterogeneity in expression of CYP3A4 and CYP3A5 in small bowel. Lack of prediction by the erythromycin breath test. Drug Metab Dispos. 1994;22(6):947-955.
- Lundahl J, Regårdh CG, Edgar B, Johnsson G. Relationship between time of intake of grapefruit juice and its effect on pharmacokinetics and pharmacodynamics of felodipine in healthy subjects. Eur J Clin Pharmacol. 1995;49(1-2):61-67.
- Guo LQ, Yamazoe Y. Inhibition of cytochrome P450 by furanocoumarins in grapefruit juice and herbal medicines. Acta Pharmacol Sin. 2004;25(2):129-136.
- Berteina-Raboin S. Flavonoids and Furanocoumarins Involved in Drug Interactions. Molecules. 2025;30(8):1676.
- Edwards DJ, Bernier SM. Naringin and naringenin are not the primary CYP3A inhibitors in grapefruit juice. Life Sci. 1996;59(13):1025-1030.
- Fukuda K, Ohta T, Yamazoe Y. Grapefruit component interacting with rat and human P450 CYP3A: possible involvement of non-flavonoid components in drug interaction. Biol Pharm Bull. 1997;20(5):560-564.
- Guo LQ, Fukuda K, Ohta T, Yamazoe Y. Role of furanocoumarin derivatives on grapefruit juice-mediated inhibition of human CYP3A activity. Drug Metab Dispos. 2000;28(7):766-771.
- Fujita K, Hidaka M, Takamura N, et al. Inhibitory effects of citrus fruits on cytochrome P450 3A (CYP3A) activity in humans. Biol Pharm Bull. 2003;26(9):1371-1373.
- Bailey DG, Dresser GK, Leake BF, Kim RB. Naringin is a major and selective clinical inhibitor of organic anion-transporting polypeptide 1A2 (OATP1A2) in grapefruit juice. Clin Pharmacol Ther. 2007;81(4):495-502.
- Bailey DG, Kreeft JH, Munoz C, Freeman DJ, Bend JR. Grapefruit juice-felodipine interaction: effect of naringin and 6′,7′-dihydroxybergamottin in humans. Clin Pharmacol Ther. 1998;64(3):248-256.
- Dresser GK, Bailey DG, Leake BF, et al. Fruit juices inhibit organic anion transporting polypeptide-mediated drug uptake to decrease the oral availability of fexofenadine. Clin Pharmacol Ther. 2002;71(1):11-20.
- Shirasaka Y, Shichiri M, Mori T, Nakanishi T, Tamai I. Major active components in grapefruit, orange, and apple juices responsible for OATP2B1-mediated drug interactions. J Pharm Sci. 2013;102(1):280-288.
- Misaka S, Yatabe J, Müller F, et al. Green tea ingestion greatly reduces plasma concentrations of nadolol in healthy subjects. Clin Pharmacol Ther. 2014;95(4):432-438.
- Abe O, Ono T, Sato H, et al. Role of (-)-epigallocatechin gallate in the pharmacokinetic interaction between nadolol and green tea in healthy volunteers. Eur J Clin Pharmacol. 2018;74(6):775-783.
- Roth M, Timmermann BN, Hagenbuch B. Interactions of green tea catechins with organic anion-transporting polypeptides. Drug Metab Dispos. 2011;39(5):920-926.
- Glaeser H, Bailey DG, Dresser GK, et al. Intestinal drug transporter expression and the impact of grapefruit juice in humans. Clin Pharmacol Ther. 2007;81(3):362-370.
- Nozawa T, Imai K, Nezu J, Tsuji A, Tamai I, Functional characterization of pH-sensitive organic anion transporting polypeptide OATP-B in human. J Pharmacol Exp Ther. 2004;308(2):438-445.
- Misaka S, Ono Y, Taudte RV, et al. Exposure of fexofenadine, but not pseudoephedrine, is markedly decreased by green tea extract in healthy volunteers. Clin Pharmacol Ther. 2022;112(3):627-634.
- Kato Y, Miyazaki T, Kano T, Sugiura T, Kubo Y, Tsuji A. Involvement of influx and efflux transport systems in gastrointestinal absorption of celiprolol. J Pharm Sci. 2009;98(7):2529-2539.
- Ieiri I, Doi Y, Maeda K, et al. Microdosing clinical study: pharmacokinetic, pharmacogenomic (SLCO2B1), and interaction (grapefruit juice) profiles of celiprolol following the oral microdose and therapeutic dose. J Clin Pharmacol. 2012;52(7):1078-1089.
- Rebello S, Zhao S, Hariry S, et al. Intestinal OATP1A2 inhibition as a potential mechanism for the effect of grapefruit juice on aliskiren pharmacokinetics in healthy subjects. Eur J Clin Pharmacol. 2012;68(5):697-708.
- Vaidyanathan S, Camenisch G, Schuetz H, et al. Pharmacokinetics of the oral direct renin inhibitor aliskiren in combination with digoxin, atorvastatin, and ketoconazole in healthy subjects: the role of P-glycoprotein in the disposition of aliskiren. J Clin Pharmacol. 2008;48(11):1323-1338.
- Sonoda J, Ogata K, Yoshikawa N, Sato K, Ikeda R, Shimodozono Y, Impact of green tea intake on the pharmacokinetics of celiprolol in healthy subjects. Int J Clin Pharmacol Ther. 2021;59(3):198-201.
- Kobayashi D, Nozawa T, Imai K, Nezu J, Tsuji A, Tamai I. Involvement of human organic anion transporting polypeptide OATP-B (SLC21A9) in pH-dependent transport across intestinal apical membrane. J Pharmacol Exp Ther. 2003;306(2):703-708.
- Dolton MJ, Roufogalis BD, McLachlan AJ. Fruit juices as perpetrators of drug interactions: the role of organic anion-transporting polypeptides. Clin Pharmacol Ther. 2012;92(5):622-630.
- Lilja JJ, Kivistö KT, Neuvonen PJ. Duration of effect of grapefruit juice on the pharmacokinetics of the CYP3A4 substrate simvastatin. Clin Pharmacol Ther. 2000;68(4):384-390.
- U.S. Food and Drug administration, Guidance for Industry Drug Interaction Studies — Study Design, Data Analysis, Implications for Dosing, and Labeling Recommendations. Silver Spring, MD, 2012.
- European Medicines Agency, Guideline on the investigation of drug interactions. London, 2012.
- Pharmaceuticals and Medical Devices Agency, Guideline on drug interaction for drug development and appropriate provision of information. Tokyo, Japan, 2019.
- Yu J, Zhou Z, Tay-Sontheimer J, Levy RH, Ragueneau-Majlessi I. Intestinal drug interactions mediated by OATPs: A systematic review of preclinical and clinical findings. J Pharm Sci. 2017;106(9):2312-2325.
- Linde K, Berner MM, Kriston L. St John’s wort for major depression. Cochrane Database Syst Rev. 2008;2008(4):CD000448.
- European Medicines Agency, Committee on Herbal Medicinal Products (HMPC). Community herbal monograph on Hypericum perforatum L., herba. London, 2008.
- European Medicines Evaluation Agency, Public statement on the risk of drug interactions with Hypericum perforatum (St John’s wort) and antiretroviral medicinal products. London, 2000.
- Scholz I, Liakoni E, Hammann F, et al. Effects of Hypericum perforatum (St John’s wort) on the pharmacokinetics and pharmacodynamics of rivaroxaban in humans. Br J Clin Pharmacol. 2021;87(3):1466-1474.
- M Mannel, Drug interactions with St John’s wort: mechanisms and clinical implications. Drug Saf. 2004;27(11):773-797.
- Rahimi R, Abdollahi M. An update on the ability of St. John’s wort to affect the metabolism of other drugs. Expert Opin Drug Metab Toxicol. 2012;8(6):691-708.
- Mai I, Störmer E, Bauer S, Krüger H, Budde K, Roots I. Impact of St John’s wort treatment on the pharmacokinetics of tacrolimus and mycophenolic acid in renal transplant patients. Nephrol Dial Transplant. 2003;18(4):819-822.
- Christians U, Strohmeyer S, Kownatzki R, et al. Investigations on the metabolic pathways of cyclosporine: I. Excretion of cyclosporine and its metabolites in human bile–isolation of 12 new cyclosporine metabolites. Xenobiotica. 1991;21(9):1185-1198.
- Maurer G, Lemaire M. Biotransformation and distribution in blood of cyclosporine and its metabolites. Transplant Proc. 1986;18(6 sup5):25-34.
- Kwei GY, Alvaro RF, Chen Q, et at. Disposition of ivermectin and cyclosporin A in CF-1 mice deficient in mdr1a P-glycoprotein. Drug Metab Dispos. 1999;27(5):581-587.
- Bauer S, Störmer E, Johne A, et al. Alterations in cyclosporin A pharmacokinetics and metabolism during treatment with St John’s wort in renal transplant patients. Br J Clin Pharmacol. 2003;55(2):203-211.
- Biber A, Fischer H, Römer A, Chatterjee SS. et al. Oral bioavailability of hyperforin from hypericum extracts in rats and human volunteers. Pharmacopsychiatry. 1998;31(sup1):36-43.
- Markowitz JS, Donovan JL, DeVane CL, et al. Effect of St John’s wort on drug metabolism by induction of cytochrome P450 3A4 enzyme. JAMA. 2003;290(11):1500-1500.
- Furukori H, Otani K, Yasui N, et al. Effect of carbamazepine on the single oral dose pharmacokinetics of alprazolam. Neuropsychopharmacology. 1998;18(5):364-369.
- Schmider J, Brockmöller J, Arold G, Bauer S, Roots I. Simultaneous assessment of CYP3A4 and CYP1A2 activity in vivo with alprazolam and caffeine. Pharmacogenetics. 1999;9(6):725-734.
- Dürr D, Stieger B, Kullak-Ublick GA, et al. St John’s Wort induces intestinal P-glycoprotein/MDR1 and intestinal and hepatic CYP3A4. Clin Pharmacol Ther. 2000;68(6):598-604.
- Wang Z, Hamman MA, Huang SM, Lesko LJ, Hall SD. Effect of St John’s wort on the pharmacokinetics of fexofenadine. Clin Pharmacol Ther. 2002;71(6):414-420.
- Komoroski BJ, Zhang S, Cai H, et al. Induction and inhibition of cytochromes P450 by the St. John’s wort constituent hyperforin in human hepatocyte cultures. Drug Metab Dispos. 2004;32(5):512-518.
- Yamasaki Y, Kobayashi K, Chiba K. Effect of pregnenolone 16α-carbonitrile on the expression of P-glycoprotein in the intestine, brain and liver of mice. Biol Pharm Bull. 2018;41(6):972-977.
- Ikeda K, Horie K, Inoue S, Blumberg B. Physiological functions and pathological roles of PXR. J Endocr Soc. 2025;9(9):bvaf119.
- Hennessy M, Kelleher D, Spiers JP, et al. St Johns wort increases expression of P-glycoprotein: implications for drug interactions. Br J Clin Pharmacol. 2002;53(1):75-82.
- Jiang TA. Health Benefits of Culinary Herbs and Spices. J AOAC Int. 2019;102(2):395-411.
- Lanzotti V, Bonanomi G, Scala F. What makes Allium species effective against pathogenic microbes?. Phytochemistry Reviews. 2013;12:751-772.
- Petrovska BB, Cekovska S. Extracts from the history and medical properties of garlic. Pharmacogn Rev. 2010;4(7):106-110.
- Moutia M, Habti N, Badou A. In Vitro and In Vivo Immunomodulator Activities of Allium sativum L. Evid Based Complement Alternat Med. 2018 Jun 12:2018:4984659.
- El-Saber Batiha G, Beshbishy AM, Wasef LG, et al. Chemical Constituents and Pharmacological Activities of Garlic (Allium sativum L.): A Review. Nutrients. 2020;12(3):872.
- Ansary J, Forbes-Hernández TY, Gil E, et al. Potential Health Benefit of Garlic Based on Human Intervention Studies: A Brief Overview. Antioxidants (Basel). 2020;9(7):619.
- Zhu Y, Anand R, Geng X, Ding Y. A mini review: garlic extract and vascular diseases. Neurol Res. 2018;40(6):421-425.
- Melguizo-Rodríguez L, García-Recio E, Ruiz C. Biological properties and therapeutic applications of garlic and its components. Food Funct. 2022;13(5):2415-2426.
- Steiner M, Li W. Aged garlic extract, a modulator of cardiovascular risk factors: a dose-finding study on the effects of AGE on platelet functions. J Nutr. 2001;131(3s):980S-984S.
- Rahman K, Billington D, Dietary supplementation with aged garlic extract inhibits ADP-induced platelet aggregation in humans. J Nutr. 2000;130(11):2662-2665.
- Apitz-Castro R, Badimon JJ, Badimon L, A garlic derivative, ajoene, inhibits platelet deposition on severely damaged vessel wall in an in vivo porcine experimental model. Thromb Res. 1994;75(3):243-249.
- Allison GL, Lowe GM, Rahman K, Aged garlic extract may inhibit aggregation in human platelets by suppressing calcium mobilization. J Nutr. 2006;136(3 sup):789S-792S.
- Malhotra U, Hasday M, Romanos GE, Javed F. Assessment of routine diet (garlic consumption) as a pre- and postoperative protocol in oral and maxillofacial surgical interventions: An evidence-based literature review. Nutr Health. 2020;26(2):135-139.
- Wong WW, Gabriel A, Maxwell GP, Gupta SC. Bleeding risks of herbal, homeopathic, and dietary supplements: a hidden nightmare for plastic surgeons?. Aesthet Surg J. 2012;32(3):332-346.
- Hatfield J, Saad S, Housewright C. Dietary supplements and bleeding. Proc (Bayl Univ Med Cent). 2022;35(6):802-807.
- Ge B, Zhang Z, Zuo Z. Updates on the clinical evidenced herb-warfarin interactions. Evid Based Complement Altern Med. 2014:2014:957362.
- Rahman K. Effects of garlic on platelet biochemistry and physiology. Mol Nutr Food Res. 2007;51(11):1335-1344.
- Briggs WH, Xiao H, Parkin KL, Shen C, Goldman IL. Differential inhibition of human platelet aggregation by selected Allium thiosulfinates. J Agric Food Chem. 2000;48(11):5731-5735.
- Rose KD, Croissant PD, Parliament CF, Levin MB. Spontaneous spinal epidural hematoma with associated platelet dysfunction from excessive garlic ingestion: a case report. Neurosurgery. 1990;26(5):880-2.
- German K, Kumar U, Blackford HN. Garlic and the risk of TURP bleeding. Br J Urol. 1995;76(4):518.
- Burnham BE. Garlic as a possible risk for postoperative bleeding. Plast Reconstr Surg. 1995;95(1):213.
- Shakeel M, Trinidade A, McCluney N, Clive B. Complementary and alternative medicine in epistaxis: a point worth considering during the patient’s history. Eur J Emerg Med. 2010;17(1):17-19.
- Nair A, Abdelqader B, Sureshkumar S, Katmawai-Sabbagh S. Hematuria and dietary supplements – A case report on pronounced bleeding following a minimally invasive urological intervention in a patient on long term garlic supplements. Urol Case Rep. 2023;50:102493.
- Hodges PJ, Kam PC. The peri-operative implications of herbal medicines. Anaesthesia. 2002;57(9):889-899.
- Piscitelli SC, Burstein AH, Welden N, Gallicano KD, Falloon J. The effect of garlic supplements on the pharmacokinetics of saquinavir. Clin Infect Dis. 2002;34(2):234-238.
- Hajda J, Rentsch KM, Gubler C, Steinert H, Stieger B, Fattinger K. Garlic extract induces intestinal P-glycoprotein, but exhibits no effect on intestinal and hepatic CYP3A4 in humans. Eur J Pharm Sci. 2010;41(5):729-735.
- Gallicano K, Foster B, Choudhri S. Effect of short-term administration of garlic supplements on single-dose ritonavir pharmacokinetics in healthy volunteers. Br J Clin Pharmacol. 2003;55(2):199-202.
- Bordes C, Leguelinel-Blache G, Lavigne JP, et al. Interactions between antiretroviral therapy and complementary and alternative medicine: a narrative review. Clin Microbiol Infect. 2020;26(9):1161-1170.
- Wiesner A, Skrońska M, Gawlik G, Marcinkowska M, Zagrodzki P, Paśko P. Interactions of Antiretroviral Drugs with Food, Beverages, Dietary Supplements, and Alcohol: A Systematic Review and Meta-analyses. AIDS Behav. 2023;27(5):1441-1468.
- Cloarec N, Solas C, Ladaique A. Sub-therapeutic darunavir concentration and garlic consumption; a «Mediterranean» drug-food interaction, about 2 cases. Eur J Clin Pharmacol. 2017;73(10):1331-1333.
- Berginc K, Trdan T, Trontelj J, Kristl A. HIV protease inhibitors: garlic supplements and first-pass intestinal metabolism impact on the therapeutic efficacy. Biopharm Drug Dispos. 2010;31(8-9):495-505.
- Jalloh MA, Gregory PJ, Hein D, Risoldi Cochrane Z, Rodriguez A. Dietary supplement interactions with antiretrovirals: a systematic review. Int J STD AIDS. 2017;28(1):4-15.
- World Health Organization. WHO report on the global tobacco epidemic, 2023: protect people from tobacco smoke. Geneva, World Health Organization, 2023. Available from: https://www.who.int/publications/detail-redirect/9789240077164 (Accessed March 17, 2026.)
- IHME, Global Burden of Disease (2025) – with major processing by Our World in Data. “Share of deaths attributed to smoking”. Available from: https://archive.ourworldindata.org/20260213-112438/grapher/share-deaths-smoking.html (Accessed March 17, 2026.)
- World Health Organization. Status of tobacco use in the Region. 19 May 2025. Available from: https://www.who.int/europe/news-room/fact-sheets/item/tobacco (Accessed March 17, 2026.)
- Perezhogina TA, Gnuchikh EV, Faizullin RI, et al. Investigation of