





Inhibitory Effects of Puerarin on Uterine Contractions
Inhibitory effects of puerarin on myometrium contraction during rat menstrual cycle and its underlying mechanisms
Qin Ma1, Wei Zhang1, Yu-dong Wang2, Zhi-feng Tian3, Zhong-rui Du1, Hong-fang Li1, 4
- Department of Physiology, College of Basic Medicine, Lanzhou University, Lanzhou 730000, China
- School of Pharmacy, Lanzhou University, Lanzhou 730000, China
- Function Laboratory in College of Basic Medicine, Lanzhou University, Lanzhou 730000, China
- Key Laboratory of Preclinical Study for New Drugs of Gansu Province, Lanzhou 730000, China
OPEN ACCESS
PUBLISHED: 30 September 2024
CITATION: Author1, D., Author2, R., et al., 2024. Inhibitory effects of puerarin on myometrium contraction during rat menstrual cycle and its underlying mechanisms. Medical Research Archives, [online] 12(9). https://doi.org/10.18103/mra.v12i9.5802
DOI: https://doi.org/10.18103/mra.v12i9.5802
ISSN: 2375-1924
ABSTRACT
Purpose: To observe uterine contractile patterns in various phases of the menstrual cycle, and elucidate mechanisms involved in the inhibition of puerarin on rat uterine contraction.
Methods: Uterus strips of Sprague Dawley rat were suspended in tissue chambers to measure the contractile activities of the strips.
Results: Isolated myometrial strips possessed three typical forms of contractions: spontaneous rhythmic contraction, tonic contraction, and irregular contraction, respectively appearing in varied phases of the rat menstrual cycle. Puerarin not only dose-dependently inhibited spontaneous contraction but also could dose-dependently inhibit KCl or PGF2α-induced uterus contractions. The inhibitory effects induced by puerarin in KCl-precontracted myometrial strips could be attenuated partly by β2 adrenoceptor antagonist ICI118551 but had no obvious changes prior to treatment with glibenclamide, Nw-L-nitro-arginine, ICI 182,780, tamoxifen or indomethacin. In high K+ Ca2+-free Kreb’s solution containing 0.01 mmol/L egtazic acid (EGTA), puerarin decreased the first phase contractions induced by oxytocin, acetylcholine, or prostaglandin F2α, but did not affect the second contractions induced by CaCl2, and also had no marked effect on the concentration-response curve of CaCl2.
Conclusion: The results demonstrate that uterus contraction patterns differ during varied menstrual cycle phases. Puerarin can inhibit isolated myometrium contractions, probably due to activation of β2-adrenoceptor, and partially via reduction of Ca2+ release from the sarcoplasmic reticulum.
Keywords: Puerarin; myometrium contraction; menstrual cycle; adrenergic receptor; Ca2+ release
INTRODUCTION
An adequate uterine movement can provide for gamete/embryo transportation through the utero-tubal cavities and successful embryo implantation in spontaneous or assisted reproduction. It has been reported that the non-pregnant uterus exhibits wave-like activity throughout the estrous cycle, but there are varied frequencies, intensities, and forms of myometrial contraction movements accompanying the different periods of the menstrual cycle. Moreover, the endogenous estrogen and progesterone secretions during the ovarian cycle have a very important modulating effect on uterine contractile activity, however little is known about the precise contractile characteristics of uterine smooth muscle from varied phases of the menstrual cycle. Therefore, the present study aimed to identify the relationship between uterine contractile patterns and the phases of the menstrual cycle, which will be very helpful for determining embryo receptivity, improving the pregnancy rate, understanding the etiology of infertility, and alleviating the symptoms of dysmenorrhea and endometriosis by regulating the uterine contractile activity.
Puerarin (4’-7-dihydroxy-β-D-glucosyl isoflavone), an analog of estradiol, is a C-glycoside compound extracted from Pueraria radix, which is used in Traditional Asian Medicine and as nutritious food. Its crude extracts contain rich isoflavones, such as daidzein, daidzin, puerarin, and 7-diglucoside, of which structures are very similar. Besides, a few studies have revealed that puerarin possesses many physiological and pharmacological functions, including apoptosis in colon cancer cells, binding preferential to estrogen β receptor, inhibitions of aromatase and xanthine oxidase expression and modulation of cytochrome P450 isoform activity. Puerarin also has the potential for clinical use because of its improvement in blood circulation, the prevention of cardiovascular diseases, and improvement of arrhythmia. Thus, puerarin injection has been widely used in clinical practice. Studies have also shown that puerarin has inhibitory effects on the contractility of isolated vascular smooth muscle in mice. Although the increasing interest in the effects of puerarin, there are few studies about the effects of puerarin on uterine smooth muscles. In our study, the second purpose was to observe and compare the effects of puerarin on the spontaneous and activated contractile activities of uterine smooth muscle in various phases of the estrous cycle and to elucidate the underlying mechanisms.
MATERIALS AND METHODS
DRUGS
Puerarin (PUE), erythro-DL-1-(7-Methylindan-4-aryloxy)-3-isopropylamine-2-butanol (ICI 118551), tamoxifen (TAM), Nw-nitro-L-arginine (L-NNA), oxytocin (OXY), acetylcholine (ACh), indomethacin (IND) were purchased from Sigma-Aldrich, St. Louis, MO Sigma, Chemical Co, USA; Glibenclamide (HB-419), prostaglandin F2α (PGF2α), 7α-[9-(4,4,5,5,5-Pentafluoro-pentyl sulphonyl)nonyl]oestra-1,3,5(10)-triene-3,17β-diol (ICI 182780) were obtained from Tocris Bioscience, Minneapolis, MN. OXY and ACh were dissolved in distilled water, while the other drugs were dissolved in dimethyl sulphoxide (DMSO).
ANIMAL AND TISSUE PREPARATION
All experiments were performed in strict accordance with the protocols approved by the Ethics Committee and Institutional Animal Care and Use Committee of Lanzhou University, and in accordance with guidelines from the International Association for the Study of Pain. Female Sprague Dawley rats (SD rats, n=50, provided by the experimental animal center of Lanzhou University) weighing 200-230g had free access to food and water before sacrifice. All animals were fasted overnight (12h) prior to being operated without limiting water. The experimental-consuming rat was anesthetized with 0.3% sodium pentobarbitone intramuscularly before euthanasia, and the whole uterus was quickly removed and placed in a Petri dish filled with Kreb’s solution containing the following compositions (mmol/L): NaCl 120, KCl 5.9, NaH2PO4 1.2, MgCl2 1.2, NaHCO3 15.4, CaCl2 2.5, and glucose 11.5, buffered at pH 7.4. The uterine horns were cut into three equal-length (5mm×10mm) segments, and then the muscle strips were mounted longitudinally in separate 5mL tissue chambers containing 37±0.5℃ Kreb’s solution, bubbled with 95% O2 and 5% CO2. The muscle preparations were allowed to equilibrate for 30min with a resting tension of 1.5g and the solution was changed every 20min. The tonic and phasic contractions of the uterine smooth muscles were measured with force transducers and recorded with the BL-420S experimental system of biological function (TME, China) by microcomputer.
CONTRACTILITY STUDY
Puerarin (0.1, 0.5, 1.0, 5.0, 10.0, 50.0, or 100.0 μmol/L) or the same dose of the solvent DMSO (control) was added cumulatively to the tissue chamber every 5min, the effects of different concentration puerarin on contractile activity of isolated uterine smooth muscle were observed. In some experiments, in order to determine the effect of PUE on contractile response stimulated by high K+ (40 mmol/L KCl), KCl was added into the organ chambers until there was nearly no contractile-wave, when the tonic contraction of uterine smooth muscles was induced by KCl to the maximal contraction, PUE (0.1-100.0 μmol/L) was administrated cumulatively, and the effect of PUE on KCl–induced uterine contraction was observed. In order to determine the mechanisms involved in the effects of PUE on KCl–induced myometrium contraction, uterine strips were treated with the following drugs (10-6mol/L ICI118551, 10-5mol/L HB-419, 10-5mol/L L-NNA, 10-6mol/L TAM, 10-5mol/L ICI182780, 10-5mol/L IND) for 5 to 10 min, and then the influence of PUE (0.1-100.0 μmol/L) on KCl-induced myometrium contraction was observed repeatedly. In order to determine the effect of PUE on contractile response stimulated by PGF2α, PGF2α (10-6 mol/L) was added into the organ chambers, and the uterine contractile activities were activated and enhanced, when the contractile activities were stable PUE (0.1-100.0 μmol/L) was added cumulatively, and the effect of PUE on PGF2α–induced uterine contraction was observed.
STATISTICAL ANALYSIS
All results are expressed as mean±SEM. “n” refers to the number of rats. Data were expressed as % decrease in KCl (40 mmol/L) induced contraction or basal contraction. In experiments involving concentration-response curves, the results were expressed as the percentage of control maximal contractile responses induced by 10-2 mol/L CaCl2. The maximum contractile response (Emax) and the half-maximum effective concentration value (EC50) were determined for each curve by using a non-linear least square (Graph Pad Software, San Diego, CA, USA) and presented as pD2 (pD2= -logEC50). Statistical analysis was performed by using the Student’s t-test and analysis of one-way ANOVA with the SPSS software (SPSS, Inc., Chicago, IL). P<0.05 was considered statistically significant.
RESULTS
RELATIONSHIP BETWEEN ESTROUS CYCLE AND CONTRACTILE ACTIVITIES OF THE UTERINE SMOOTH MUSCLE IN RATS
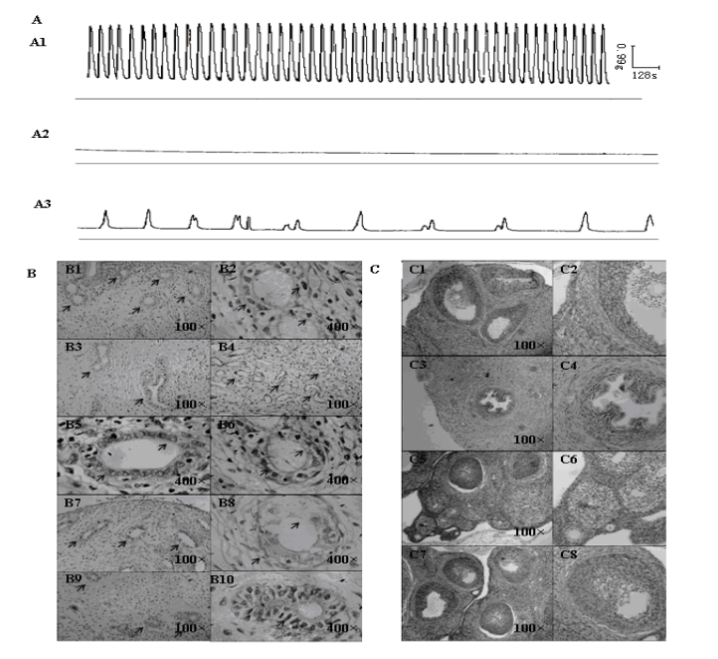
The spontaneous contractile activities of isolated uterine smooth muscle from SD rats were not always regular. There were three typical contractile patterns, which corresponded to different phases of the menstrual cycle. (1) Some isolated smooth muscle strips possessed spontaneous phasic contraction with mean basal resting tension 1.594±0.12g, mean amplitude 3.035±0.39g, and mean frequencies 0.938±0.15 min/wave (Figure 1A: A1). The rhythmic contractile waves were tight, uniform, and stable; by hematoxylin-eosin staining (HE staining) observation, the uterine cycle was in the late proliferative phase or early secretory phase (Figure 1B: B1, B2), and ovary seemed to be in the maturation phase of follicle or the ovulatory period (Figure 1C: C1, C2). The glands in the uterine tissue increased and became bigger (Figure 1B: B1), and the shape of the glands appeared near round; the shape of the follicular epithelial cell nucleus became a short rod from a long rod (Figure 1B: B2); the arrangement of the interstitial cell was neat and tight. (2) The other muscle strips had obvious tonic contraction with mean basal resting tension of 1.281±0.07g (Figure 1A: A2), there were nearly no waves in the contractile traces, and even exhibited a straight line only; according to the observation of HE staining, the uterine cycle was in the late secretory phase (Figure 1B: B3, B4, B5, B6), and ovary showed in the luteal phase (Figure 1C: C3, C4). The uterus glands became even bigger, and looked like ellipses or dumbbells (Figure 1B: B3, B4); there appeared some bubbles near the nucleus of the follicular epithelial cells (Figure 1B: B5), and some of them showed apocrine secretion (Figure 1B: B6); the interstitial cells were round; there were many spiral arteries in the tissue (Figure 1B: B4). (3) The rest of the smooth muscle strips showed untypical contractile activities (Figure 1A: A3): the waves were not rhythmic and regular, the amplitude and frequencies of the waves were not stable, and the waves became weaker and weaker, even disappeared at last; it occurred in the menstrual period (Figure 1B: B7, B8) or the early proliferative phase of the uterus (Figure 1B: B9, B10), and ovary showed in the early (Figure 1, C: C5, C6) or the mid of follicle phase through HE staining (Figure 1C: C7, C8). The uterus glands in the functional stratum became broken (Figure 1B: B7, B8), the interstitial cells tended to get together, and there appeared a small number of neutrophils; Additionally, some slices showed us a few small and round glands (Figure 1B: B9), the shape of the follicular epithelial cell nucleus looked like a long rod (Figure 1B: B10), the arrangement of the interstitial cell was very neat and tight.
EFFECTS OF PUERARIN ON SPONTANEOUS PHASIC CONTRACTIONS IN RAT UTERINE TISSUES
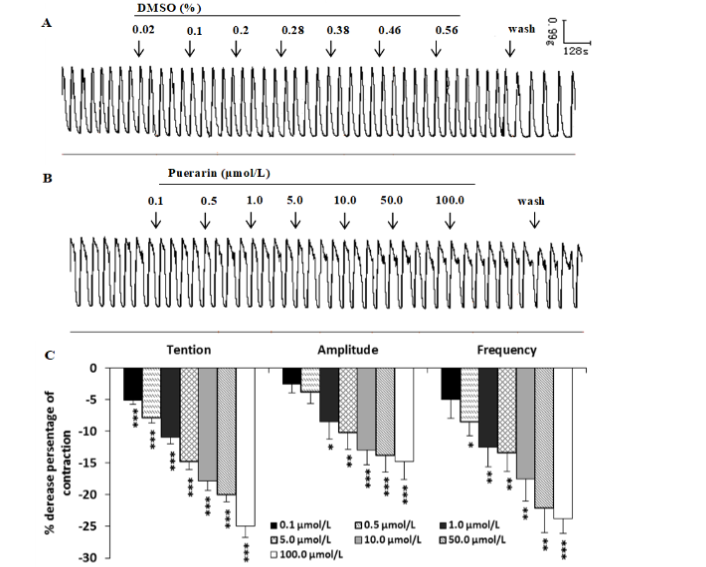
In some uterine strips with spontaneous rhythmic contractions, puerarin (0.1-100.0μmol/L) could dose-dependently inhibit the phasic contractile activities, it could decrease the mean contractile amplitude and frequencies significantly, also produced an obvious reduction in resting tone compared to the solvent DMSO control (Figure 2).
Effects of a variety of inhibitors on puerarin-induced uterine relaxation in KCl-precontracted uterine strips
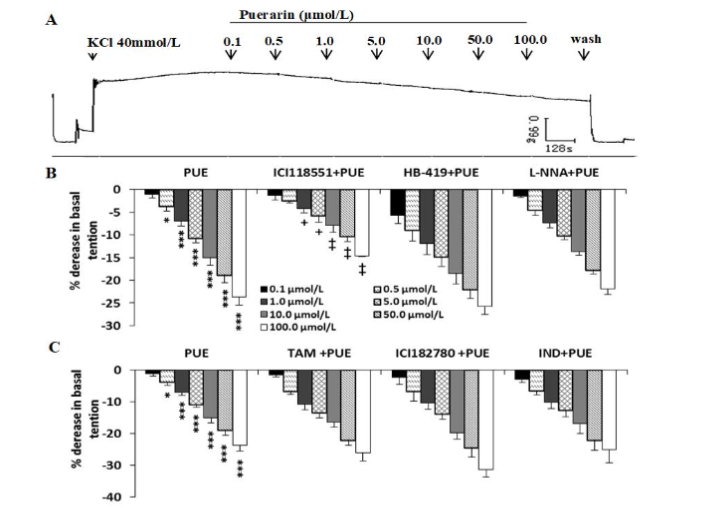
In KCl (40 mmol/L) precontracted uterine smooth muscle, puerarin (0.1-100 μmol/L) produced a dose-dependent relaxation (Figure 3A, r=0.96, P<0.05, n=10). Incubation with ICI 118551, a β2 adrenergic antagonist, partly but significantly inhibited the concentration-dependent relaxation caused by puerarin in KCl-precontracted uterine strips (n=5, Figure 3 B), however pretreatment with ATP-dependent K+ channel blocker HB-419 (n=8), NO synthase inhibitor L-NNA (n=7) (Figure 3B), estrogen receptor modulator TAM (n=9) or antagonist ICI 182780 (n=7), and prostaglandin synthase inhibitor IND (n=9) (Figure 3C) respectively, failed to alter the inhibitory effects of puerarin.
EFFECTS OF PUERARIN ON PGF2α-INDUCED UTERINE CONTRACTIONS
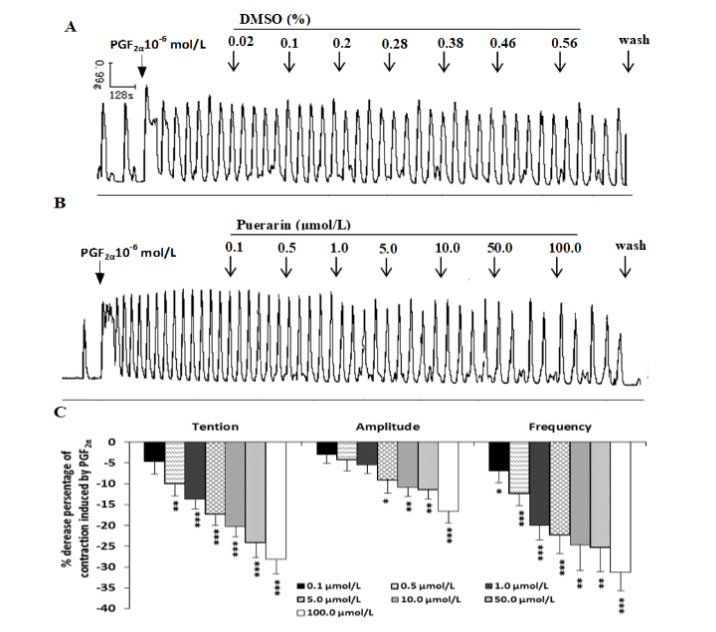
As shown in Figure 4 A, PGF2α (10-6 mol/L) increased contractile force, amplitude, and frequencies, puerarin (0.1-100.0 μmol/L) markedly inhibited the PGF2α-induced contraction in a dose-dependent manner, it could decrease the mean contractile amplitude, frequencies significantly and basal tension as compared to the solvent DMSO control (Figure 4A, B and C).
EFFECTS OF PUERARIN ON BIPHASIC CONTRACTIONS INDUCED BY OXY, ACh, PGF2α, AND CaCl2
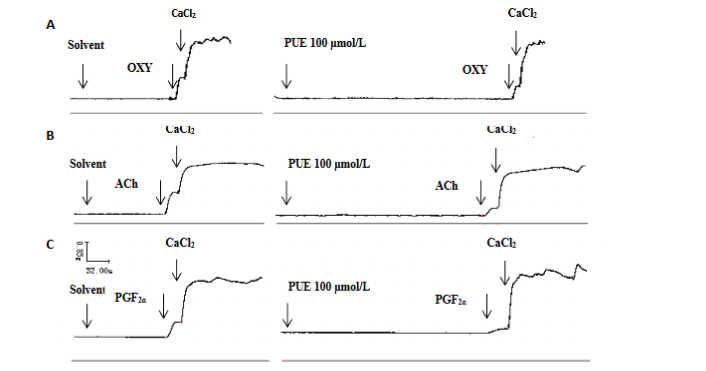
In calcium-free (0.01 mmol/L EGTA) Kreb’s solution, no spontaneous phasic contractions were observed, but OXY (0.4IU/L), ACh (5×10-5 mol/L), or PGF2α (10-5 mol/L) could cause a transient contraction with the tensive increase of 0.913±0.19g, 1.896±0.37g, 0.428±0.09g respectively. As soon as such contraction reached a plateau, CaCl2 (20 mmol/L) was rapidly added into the bath and another higher contractile response occurred with the tensive increase of 2.033±0.41g (n=12), 1.948±0.64g (n=6), 1.726±0.36g (n=6) respectively. Puerarin (100 μmol/L) reduced the first contraction induced by OXY from 0.913±0.19g to 0.522±0.09g (P<0.05, Figure 5 A), by ACh from 1.896±0.37g to 1.140±0.36g (P<0.05, Figure 5 B), and by PGF2α from 0.428±0.09g to 0.176±0.13g (P<0.05, Figure 5 C), but had no obvious changes in the second contraction caused by CaCl2 in Figure 5A (2.03±0.41g in puerarin groups vs 2.26±0.36g in solvent control, P>0.05), in Figure 5B (1.95±0.64g in puerarin groups vs 2.16±0.85g in solvent control, P>0.05), and in Figure 5C (1.73±0.36g in puerarin groups vs 1.79±0.37g in solvent control, P>0.05).
EFFECTS OF PUERARIN ON Ca2+-DEPENDENT UTERINE CONTRACTIONS IN RATS
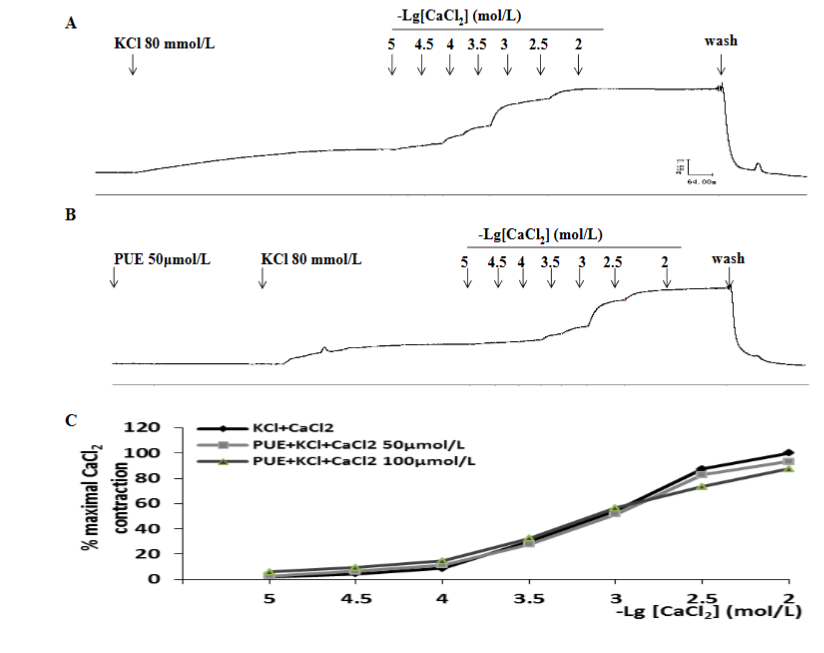
In high K+ (80mmol/L) depolarized tissues in a Ca2+-free medium, CaCl2 (10-5 to 10-2 mol/L) elicited a concentration-dependent contraction. Pretreatment with puerarin (50 or 100μmol/L) did not change the contractile response to CaCl2. The maximal contractions of CaCl2 for control and after incubation with puerarin (50 or 100μmol/L) were (100±0) %, (93.15±14.00) %, and (94.20±8.12) % respectively (P>0.05 vs control, n=9). The pD2 values of CaCl2 for control and after incubation with puerarin (50 or 100μmol/L) were 3.03±0.06, 3.03±0.10 and 3.04±0.13 respectively (P>0.05 vs control, n=9).
DISCUSSION
The non-pregnant uterus can exhibit wave-like activity, the wave frequencies can be analyzed through the unidirectional waves (fundus-to-cervix or cervix-to-fundus) in spontaneous estrous cycles and stimulation cycles. There are contraction properties and a variation in frequencies of the unidirectional waves from the mid-follicular to the late follicular phase or the early luteal phase. After ovulation, there is a reduction which might optimize the contact between the blastocyst and the endometrium to facilitate implantation. During the luteal phase, the upper fundal part of the uterus shows relative quiescence that could facilitate embryo implantation. In our present study, we observed similar results which were consistent with previous research and papers. In the late proliferative phase or early secretory phase, the contractile activities of uterine smooth muscle revealed the pattern of spontaneous phasic contraction; in the late secretory phase, there were nearly no contracting waves; and in the menstrual period and early proliferative phase, there were untypical contractile activities. These three patterns of uterine contractile activity circulated in various phases of the estrous cycle.
Steroid hormones estradiol and progesterone exert actions on uterine contractility, and they can influence the rhythmicity of uterine contractions. The endogenous estrogen secretion is responsible for the change in direction of contractions during estrus; while progesterone can cause local vasodilatation and induce uterine musculature quiescence by promoting nitric oxide synthesis in the decidua. In the present study, there appeared three different uterine contractile patterns according to the estrous cycle. All the above substantial pieces of evidence suggest that the appearance of contractile patterns could be adjusted by the steroid hormone whose secretion is fluctuated in the estrous cycle. Adequate uterine contractility may provide for gamete/embryo transportation and successful spontaneous embryo implantation or assisted reproduction, but inadequate uterine contractility may lead to ectopic pregnancies, miscarriages, retrograde bleeding with dysmenorrhea, and endometriosis. Therefore, it is possible to correct the direction of sperm movement and improve the rate of embryo implantation by adjusting the secretion and concentration of estradiol and progesterone; moreover, the steroid hormones could potentially alleviate the symptoms of dysmenorrhea and endometriosis by regulating the uterine contractile activity.
Previous studies have demonstrated that the increase of Ca2+ concentration in the uterine smooth muscle induces its contraction and the concentration of Ca2+ can be regulated by two different Ca2+ channels: receptor-operated channels (ROCs) and potential dependent calcium channels (PDCs). In the PDCs, membrane depolarization caused by high K+ (high concentrations of KCl) could open the L-type PDCs and increase the Ca2+ influx through the PDCs, resulting in uterine smooth muscle contraction. It has also been reported that potassium-stimulated uterine contraction depends exclusively upon the influx of extracellular calcium. In the present experiment, KCl (40mmol/L) could cause the tonic contraction in uterine smooth muscle strips, and puerarin could dose-dependently reduce the KCl-induced contraction in normal Kreb’s solution. The results indicate that puerarin can inhibit the influx of extracellular Ca2+, and decrease the tonic contractile activity of the myometrium. However, in our study, puerarin (50 or 100μmol/L) could not reduce CaCl2-induced contraction in a calcium-free high K+ depolarization medium and did not affect CaCl2 cumulative concentration-response curves. Considering these observations mentioned above, it seems reasonable to suggest that the mechanism of extracellular Ca2+ influx from PDCs may not be involved in the inhibitory effects of puerarin. Additionally, OXY, ACh, and PGF2α can induce uterine contractions when they bind to membrane G-protein-coupled receptors, and they increase the concentration of Ca2+ via both the influx of extracellular Ca2+ through ROCs and by the release of intracellular stored Ca2+. In the present study, OXY, ACh, or PGF2α caused a transient contraction of the uterine smooth muscle in the Ca2+-free Kreb’s solution, which involved in the release of intracellular stored Ca2+, then the addition of CaCl2 to the perfusion caused another brief contraction which induced by the influx of extracellular Ca2+ through ROCs; it is interesting that puerarin could reduce the first contraction induced by OXY, ACh, or PGF2α, but did not change the second contraction caused by CaCl2. The results suggest that the inhibitory effect of puerarin is related to the Ca2+ release from the sarcoplasmic reticulum, but may have nothing to do with the Ca2+ influx.
ICI 118,551 possesses a high degree of selectivity and specificity for the β2-adrenoceptor. By binding to cardiac β-adrenoceptor, ICI 118,551 can block the binding (and therefore action) of the endogenous catecholamines such as adrenaline and noradrenaline, resulting in a reduction in the rate and force of cardiac contraction. Thus, ICI 118,551, as a very selective β2 adrenergic receptor antagonist, was used in the present experiment to assess whether the inhibitory effects of puerarin on the uterine contractile activity were relevant to the β2-adrenoceptor, and the data from the present study suggest that the inhibitory effect of puerarin on the uterine contractile activity probably has the relationship with the β2-adrenoceptor because ICI 118,551 could partly block the inhibitory effect of puerarin.
Papers have previously shown that glibenclamide (HB-419) is a selective antagonist of the K+ channel openers in vitro. HB-419 has been demonstrated to inhibit the opening of ATP-dependent K+ channels in the pancreas, the heart and vascular smooth muscle cells. In the present study, the inhibitory effects of puerarin were not abolished by HB-419 in the KCl-induced uterine smooth muscle, it suggests that the effects of puerarin are not mediated through the activation of ATP-dependent K+ channels in uterine smooth muscle cells.
Nitric oxide (NO), previously identified as the main endothelium-derived relaxing factor, is formed from L-arginine by NO synthase (NOS) which converts L-arginine into citrulline and NO in the presence of NADPH and other cofactors. The major mechanism of NO action in most tissues is that NO can activate the soluble guanylate cyclase, and form cGMP with subsequent. In the present study, the inhibitory effects of puerarin on KCl-precontracted uterine smooth muscles were not changed by L-NNA, the inhibitor of the NO synthase, it suggests that the effects of puerarin are not relative to the NO production. Many studies prompted to assess the role of the smooth muscle relaxing factor NO in the regulation of uterine contractility and suggest that NO donors are in fact capable of altering uterine contractility, either by stimulating contractions or by inhibiting spontaneous contractions. All these pieces of evidence prove that NO is supposed to change uterine contractility; but in the present study, L-NNA could not change the effects of puerarin, which means that the effect of puerarin is not relevant to NO.
The uterus is considered a very important target organ expressed with functional estrogen receptors (ERs), so the effect of phytoestrogen puerarin may exert its estrogen effects in combination with ERs. The results of our study, however, demonstrate that the inhibitory action induced by puerarin is unlikely to be mediated by the ERs because ER modulator TAM or antagonist ICI 182780 can block not only classical ERα but also the novel ERβ failed to affect the inhibitory effects induced by puerarin, but whether these inhibitory effects of puerarin are independent of gene-mediated events for the molecular weight of puerarin is not very large, and it can easily enter cytoplasm through the cellular membrane to affect the expression of some genes, which still need more data to be proved.
In our present study, indomethacin did not change the inhibitory effect induced by puerarin on the isolated uterine smooth muscle strips from the rats, and as we also know, indomethacin is an inhibitor of the endogenous prostaglandin synthesis, all these pieces of evidence indicate that the endogenous prostaglandin synthesis is not involved in the inhibitory effects on the uterine contractile activity induced by puerarin.
Prostaglandin F2α (PGF2α) could cause the constriction of uterine smooth muscle and small endometrial blood vessels after tissue ischemia and endometrial disintegration, resulting in bleeding and pain. It has been previously reported that PGF2α levels would be elevated in women with primary dysmenorrhea. Besides, dysmenorrhea leads to increased PG (PGE2 and PGF2α) production, which may result in contraction of the blood vessels and myometrium, and insufficient blood flow to the endometrium, which in turn causes ischemia and the pain symptoms associated with dysmenorrhea. Though the role of PGs is implicated in dysmenorrhea, it is considered that dysmenorrhea is directly related to elevating PGF2α levels, and it is well established that PGF2α increases the concentration of Ca2+ and then stimulates uterine contraction. There is still a paper showing that estrogen administration could irritate the endometrial waves or contractions, but from the results and data of our laboratory in recent years, estradiol cannot stimulate the contraction of various smooth muscles, but inhibit their contractions, thus, it is reasonable to presume that the symptom of dysmenorrhea is not caused by the secretion and concentration of estradiol, but caused by the out-of-balance of the concentration between estradiol and PGF2α. The present study shows that puerarin could suppress PGF2α-induced uterine contractions of uterine smooth muscles. In Western medicine, dysmenorrhea can be treated by nonsteroidal anti-inflammatory drugs, which can produce many side effects, so Chinese medicinal therapy is considered a feasible alternative for treating dysmenorrhea. In the present study, puerarin could suppress PGF2α-induced uterine contractions of uterine smooth muscles, which suggests that the administration of puerarin might be beneficial for treating or improving dysmenorrhea.
In summary, our findings suggest that the inhibitory effects of puerarin are probably due to the activation of β2-adrenoceptor and the inhibition of Ca2+ release from the sarcoplasmic reticulum; but are not relative to the Ca2+ influx through PDCs and ROCs, the activation of ATP-dependent K+ channels, NO release, estrogen receptor, or the endogenous prostaglandin synthesis.
ACKNOWLEDGMENTS
We thank members of the functional laboratory for basic equipment during the process of this work, and Wang CH for sections of rat uterus.
SOURCES OF FUNDING
This work was supported by the Natural Science Foundation of Gansu province (20JR5RA298) and the Innovation Group of Science and Technology Project of Gansu Province (20JR5RA310).
AUTHOR CONTRIBUTIONS
HL designed the experiment and wrote the paper. QM, WZ, and ZD performed the experiments. QM and YW analyzed the data. YW and ZT contributed reagents/materials/analysis tools. QM drafted the paper. HL, YW revised the final paper. The authors read and approved the final manuscript.
DECLARATIONS
CONFLICT OF INTEREST: The authors declare that they have no conflict of interest.
ETHICAL APPROVAL: All experiments were performed in strict accordance with the protocols approved by the Ethics Committee and Institutional Animal Care and Use Committee of Lanzhou University, and in accordance with guidelines from International Association for the Study of Pain.
REFERENCES
- de Alencar Silva A, Pereira-de-Morais L, Rodrigues da Silva RE, de Menezes Dantas D, Brito Milfont CG, Gomes MF, et al. Pharmacological screening of the phenolic compound caffeic acid using rata orta, uterus and ileum smooth muscle. Chem Biol Interact. 2020;332: 109269. doi: 10.1016/j.cbi.2020.109269
- Domino M, Pawlinski B, Gajewska M, Jasinski T, Sady M, Gajewski Z. Uterine EMG activity in the non-pregnant sow during estrous cycle. BMC Vet Res. 2018;14(1):176. doi: 10.1186/s12917-018-1495-z
- Dodds KN, Staikopoulos V, Beckett EA. Uterine Contractility in the Nonpregnant Mouse: Changes During the Estrous Cycle and Effects of Chloride Channel Blockade. Biol Reprod. 2015;92(6):141. doi: 10.1095/biolreprod.115.129809
- Oki T, Douchi T, Maruta K, Nakamura S, Nagata Y. Changes in endometrial wave-like movements in accordance with the phases of menstrual cycle. J Obstet Gynaecol Res. 2002;28(3):176-181. doi: 10.1046/j.1341-8076.2002.00026.x
- Yi Y, Adrjan B, Li J, Hu B, Roszak S. NMR studies of daidzein and puerarin: active anti-oxidants in traditional Chinese medicine. J Mol Model. 2019;26;25(7):202. doi: 10.1007/s00894-019-4090-8
- Udomsin O, Krittanai S, Kitisripanya T, Tanaka H, Putalun W. A New Highly Selective and Specific Anti-puerarinpolyclonal Antibody for Determination of Puerarin Using a Mannich Reaction Hapten Conjugate. Pharmacogn Mag.2018;13(Suppl 4): S845-S851. doi: 10.4103/pm.pm_276_17
- Zhou DD, Luo M, Shang A, Mao QQ, Li BY, Gan RY, et al. Antioxidant Food Components for the Prevention and Treatment of Cardiovascular Diseases: Effects, Mechanisms, and Clinical Studies. Oxid Med Cell Longev. 2021;2021:6627355. doi: 10.1155/2021/6627355
- Zhang XL, Wang BB, Mo JS. Puerarin 6″-O-xyloside possesses significant antitumor activities on colon cancer through inducing apoptosis. Oncol Lett. 2018;16(5):5557-5564. doi: 10.3892/ol.2018.9364
- Gray SL, Lackey BR, Boone WR. Impact of kudzu and puerarin on sperm function. Reprod Toxicol. 2015;53:54-62. doi: 10.1016/j.reprotox.2015.03.01
- Yu J, Zhao L, Zhang D, Zhai D, Shen W, Bai L, et al. The effects and possible mechanisms of puerarin to treat endometriosis model rats. Evid Based Complement Alternat Med. 2015;2015:269138. doi: 10.1155/2015/269138
- Zhao CP, Chen GY, Wang Y, Chen H, Yu JW, Yang FQ. Evaluation of Enzyme Inhibitory Activity of flavonoids by Polydopamine-Modified Hollow Fiber-Immobilized Xanthine Oxidase. Molecules. 2021;26(13):3931. doi: 10.3390/molecules26133931
- Guo YJ, Liang DL, Xu ZS, Ye Q. In vivo inhibitory effects of puerarin on selected rat cytochrome P450 isoenzymes. Pharmazie. 2014;69(5):367-370
- Ma J, Wang C, Sun Y, Pang L, Zhu S, Liu Y, et al. Comparative study of oral and intranasal puerarin for prevention of brain injury induced by acute high-altitude hypoxia. Int J Pharm. 2020;15:591:120002. doi: 10.1016/j.ijpharm.2020.120002
- Zhou YX, Zhang H, Peng C. Effects of puerarin on the Prevention and Treatment of Cardiovascular Diseases. Front Pharmacol. 2021;12:771793. doi: 10.3389/fphar.2021.771793
- Jiang Z, Cui X, Qu P, Shang C, Xiang M, Wang J. Roles and mechanisms of puerarin on cardiovascular disease: A review. Biomed Pharmacother. 2022;147:112655. doi: 10.1016/j.biopha.2022
- Lin YH, Ni XB, Zhang JW, Ou CW, He XQ, Dai WJ, et al. Effect of puerarin on action potential and sodium channel activation in human hypertrophic cardiomyocytes. Biosci Rep. 2022;28;40(2):BSR20193369. doi: 10.1042/BSR20193369
- Chen M, Xiang L, Wu G, Liao Y, Cai Y. Puerarin Inhibits Endothelium-Dependent Contractions in Mouse Carotid Arteries. Med Sci Monit. 2022;26:e923163. doi: 10.12659/MSM.923163
- Domino M, Domino K, Pawlinski B, Sady M, Gajewska M, Gajewski Z. Computational multivariate modeling of electrical activity of the porcine uterus during spontaneous and hormone-induced oestrus. Exp Physiol. 2019;104(3):322-333. doi: 10.1113/EP087451
- Modzelewska B, Jóźwik M, Jóźwik M, Tylicka M, Kleszczewski T. The effects of extended nitric oxide release on responses of the human non-pregnant myometrium to endothelin-1 or vasopressin. Pharmacol Rep. 2019;71(5):892-898. doi: 10.1016/j.pharep.2019.05.003
- Hutchings G, Williams O, Cretoiu D, Ciontea SM. Myometrial interstitial cells and the coordination of myometrial contractility. J Cell Mol Med. 2009;13(10):4268-4282. doi: 10.1111/j.1582-4934.2009.00894.x
- Wahabi HA, Fayed AA, Esmaeil SA, Bahkali KH. Progestogen for treating threatened miscarriage. Cochrane Database Syst Rev. 2018;8(8):CD005943. doi: 10.1002/14651858
- Mirdamadi M, Kothencz A, Szűcs E, Benyhe S, Szécsi M, Gáspár R. Non-genomic actions of sex hormones on pregnant uterine contractility in rats: An in vitro study at term. Life Sci. 2020;263:118584. doi: 10.1016/j.lfs.2020.118584
- An SM, Kim SS, Kim J, Park MN, Lee JE, Cho SK, et al. Expression of reproductive hormone receptors and contraction-associated genes in porcine uterus during the estrous cycle. Mol Med Rep. 2017;15(6):4176-4184. doi: 10.3892/mmr.2017.6518
- MacLean JA 2nd, Hayashi K. Progesterone Actions and Resistance in Gynecological Disorders. Cells. 2022;11(4):647. doi: 10.3390/cells11040647
- Liu J, Feng R, Dai O, Ni H, Liu LS, Shu HZ, et al. Isoindolines and phthalides from the rhizomes of Ligusticum chuanxiong and their relaxant effects on the uterine smooth muscle. Phytochemistry. 2022;198:113159. doi: 10.1016/j.phytochem.2022.113159
- Ali A, Redfors B, Lundgren J, Alkhoury J, Oras J, Gan LM, et al. Effects of pretreatment with cardio stimulants and beta-blockers on isoprenaline-induced takotsubo-like cardiac dysfunction in rats. Int J Cardiol. 2019;15:281:99-104. doi: 10.1016/j.ijcard.2018.12.045
- Bhatiya S, Choudhury S, Gari M, Singh P, Shukla A, Garg SK. Myometrial Calcium and Potassium Channels Play a Pivotal Role in Chromium-Induced Relaxation in Rat Uterus: an In Vitro Study. Biol Trace Elem Res. 2020;198(1):198-205. doi: 10.1007/s12011-020-12040-1
- Passaglia P, Gonzaga NA, Tirapelli DP, Tirapelli LF, Tirapelli CR. Pharmacological characterization of the mechanisms underlying the relaxant effect of adrenomedullin in the rat carotid artery. J Pharm Pharmacol. 2014;66(12):1734-1746. doi: 10.1111/jphp.12299
- Babes A, Fischer MJ, Filipovic M, Engel MA, Flonta ML, Reeh PW. The anti-diabetic drug glibenclamide is an agonist of the transient receptor potential Ankyrin 1 (TRPA1) ion channel. Eur J Pharmacol. 2013;04(1-3):15-22. doi: 10.1016/j.ejphar.2013.02.018
- Negroni JA, Lascano EC, del Valle HF. Glibenclamide action on myocardial function and arrhythmia incidence in the healthy and diabetic heart. Cardiovasc Hematol Agents Med Chem. 2007;5(1):43-53. doi: 10.2174/187152507779315868
- Zhang R, Zou Z, Zhou X, Shen X, Fan Z, Xie T, et al. Comparative Effects of Three Sulfonylureas (Glibenclamide, Glimepiride, and Gliclazide) on Proliferation and Migration of Vascular Smooth Muscle Cells. Cell Physiol Biochem. 2019;52(1):16-26. doi: 10.33594/000000002
- Förstermann U, Sessa WC. Nitric oxide synthases Eur Heart J. 2012;33(7):829-837, 837a-837d. doi: 10.1093/eurheartj/ehr304
- Picciano AL, Crane BR. A nitric oxide synthase-like protein from Synechococcus produces NO/NO3 from L-arginine and NADPH in a tetrahydrobiopterin- and Ca2+-dependent manner. J Biol Chem. 2019;294(27):10708-10719. doi: 10.1074/jbc.RA119.008399
- Totzeck M, Hendgen-Cotta UB, Luedike P, Berenbrink M, Klare JP, Steinhoff HJ, et al. Nitrite regulates hypoxic vasodilation via myoglobin-dependent nitric oxide generation. Circulation. 2012;126(3):325-334. doi: 10.1161/CIRCULATIONAHA.111.087155
- Khan FA, Chenier TS, Murrant CL, Foster RA, Hewson J, Scholtz EL. Dose-dependent inhibition of uterine contractility by nitric oxide: A potential mechanism underlying persistent breeding-induced endometritis in the mare. Theriogenology. 2017;90:59-64. doi: 10.1016/j.theriogenology.2016.11.026
- Yallampalli C, Izumi H, Byam-Smith M, Garfield RE. An L-arginine-nitric oxide-cyclic guanosine monophosphate system exists in the uterus and inhibits contractility during pregnancy. Am J Obstet Gynecol. 1994;170(1 Pt 1):175-185. doi: 10.1016/s0002-9378(94)70405-8
- Powell SA, Smith BB, Timm KI, Menino AR. Expression of estrogen receptors alpha and beta in the corpus luteum and uterus from non-pregnant and pregnant llamas. Mol Reprod Dev. 2007;74(8):1043-1052. doi: 10.1002/mrd.20684
- Nathan MR, Schmid P. A Review of Fulvestrant in Breast Cancer. Oncol Ther. 2017;5(1):17-29. doi: 10.1007/s40487-017-0046-2
- Dawood MY, Khan-Dawood FS. Differential suppression of menstrual fluid prostaglandin F2α, prostaglandin E2, 6-keto prostaglandin F1α and thromboxane B2 by suprofen in women with primary dysmenorrhea. Prostaglandins Other Lipid Mediat. 2007;83(1-2):146-153. doi: 10.1016/j.prostaglandins.2006.10.009
- Gu B, Zhu S, Ding X, Deng Y, Ma X, Gan J, et al. Effect of Low-Power Visible-Light-Activated Photodynamic Therapy (PDT) on primary dysmenorrhea: A Multicenter, Randomized, Double-Blind, Placebo-Controlled Trial. Int J Womens. 2022;14:1029-1036
- Yang W, Guo H, Niu J, Liu J, Su R, Bai Y, et al. Phloroglucinol inhibits oxytocin-induced contraction in rat gastric circular muscle and uterine smooth muscle. J Obstet Gynaecol. 2023;43(1):2130208. doi: 10.1080/01443615.2022.2130208
- Earl RA, Grivell RM. Nifedipine for primary dysmenorrhoea. Cochrane Database Syst Rev. 2021;12(12):CD012912. doi: 10.1002/14651858
- Sun L, Liu L, Zong S, Wang Z, Zhou J, Xu Z, et al. Traditional Chinese medicine Guizhi Fuling capsule used for therapy of dysmenorrhea via attenuating uterus contraction. J Ethnopharmacol. 2016;191:273-279. doi: 10.1016/j.jep.2016.06.042
- Gestel IV, lJland MM, Hoogland HJ, Evers JLH. Endometrial wave-like activity in the non-pregnant uterus. Human Reproduction Update. 2003;9(2):131-138. doi: 10.1093/humupd/dmg011
- Li G, Liu A, Lin M, Liao S, Wen Z. Chinese herbal formula siwutang for treating primary dysmenorrhea: A systematic review and meta-analysis of randomized controlled trials. Maturitas. 2020;138:26-35. doi: 10.1016/j.maturitas.2020.03.009