





T Cell Repertoire and Autoreactivity in Lupus SLE
T Cell Repertoire in Lupus: Autoreactive T Cells in Central and Peripheral Selection
Sun Jung Kim1, Betty Diamond2
- Center for Autoimmune, Musculoskeletal and Hematopoietic Disease, The Feinstein Institute for Medical Research, Manhasset, NY 11030
- Department of Molecular Medicine, Northwell Health-Hofstra School of Medicine, Hofstra University, Hempstead, NY 11549
OPEN ACCESS
PUBLISHED: 31 August 2024
CITATION: Kim, DS. and Diamond, B., 2024. T Cell Repertoire in Lupus: Autoreactive T Cells in Central and Peripheral Selection. Medical Research Archives, [online] 12(8). https://doi.org/10.18103/mra.v12i8.5637
COPYRIGHT: © 2025 European Society of Medicine. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
DOI https://doi.org/10.18103/mra.v12i8.5637
ISSN 2375-1924
ABSTRACT
Systemic lupus erythematosus (SLE) is an immune disorder characterized by the presence of T and B cells which drive a pathogenic autoimmune response. Autoreactive T effector cells naturally arise during the process of development in the thymus, but most are removed by several self-tolerance mechanisms. Self-antigen recognizing T cells emigrating from the thymus are normally prevented from causing cellular or tissue damage by peripheral tolerance mechanisms. While we understand much about how self-antigens are presented to T cells and the mechanisms to remove those T cells or convert them into regulatory T cells in thymus and peripheral lymphoid organs, we understand much less about the derivation of potentially pathogenic self-reactive T cells in autoimmune disease. It is not clear whether individuals with autoreactive T cell activation have an altered T cell receptor repertoire of naïve T cells that have matured to immunocompetence as effector cells or whether the pathogenic process involves altered activation of immunocompetent T cells in the periphery. There is increasing evidence that the autoantibodies that characterize SLE result from an interaction of B cells with self-reactive T cells. Here we summarize the evidence for autoreactive T cells in both lupus mouse models and in patients with SLE and discuss the evidence for either altered T cell selection or altered T cell activation in SLE, which has implications for both pathogenesis and treatment.
Keywords
Systemic lupus erythematosus, autoreactive T cells, T cell receptor repertoire, immune tolerance, autoantibodies
Introduction
The T cell receptor (TCR) is generated by combining gene segments by V(D)J recombination which leads to a diverse repertoire of TCRs (reviewed in 1). Mature T cells express the α and β chain of the TCR, each comprising of variable (V) and constant (C) regions. The C region of each chain is identical, but the V region contains the hypervariable complementary-determining region 3 (CDR3) which directly contacts peptides and CDR1 and CDR2 loops which contact the MHC helices (reviewed 2) to allow the TCR to bind a peptide-MHC complex (p-MHC). The selection of the repertoire of naïve T cells mainly occurs during T cell development in the thymus through both a positive and a negative selection process. Recognition of a p-MHC by a TCR is the first fate-determining step for antigen-specific T cells. Selection of antigen-reactive T cells is further refined when the naïve T cells encounter peptide-presenting antigen presenting cells (APCs) in the periphery. The repertoire of CD4+ follicular helper T (Tfh) cells is critical to the humoral immune response since many B cell responses require T help through a cognate interaction between p-MHC II on APCs and the TCR of the Tfh cell. Tfh cells are important in the pathogenesis of SLE as they promote autoantibody production by B cells (reviewed in 3). The signal which is initiated by the p-MHC II on the B cell and the TCR on the T cell determines the quality and quantity of B cell activation and differentiation in a germinal center (GC) response which generates high affinity antibodies and B cell memory.
Systemic lupus erythematosus (SLE) is a complex disease characterized by dysregulation of several immune cell lineages. Pathogenic autoantibodies form pro-inflammatory immune complexes or directly bind and damage tissues. T cells are integral to disease initiation and pathogenesis as they produce inflammatory cytokines and provide help for autoreactive B cells (reviewed in 3 4). Mechanisms of T cell selection and T cell tolerance in the thymus are essential to the generation of the T cell repertoire and the function of APCs is crucial to the mechanism of tolerance and activation in the periphery. The defect in the regulation of autoreactive T cells in SLE is not understood, although murine models suggest that disruption of peripheral mechanisms of tolerance are important in this disease.
T cells reactive with several self-antigens targeted by autoantibodies have been identified in individuals with SLE. B cells reacting with nuclear antigens are censored during B cell maturation in both healthy individuals and patients with SLE 5, but some autoreactive B cells remain in the system. Those autoreactive B cells, however, stay inactive (anergy) or quiescent unless they receive activation signals from interacting Th cells. While B cells can produce antibodies in a T-independent manner, production of many pathogenic autoantibodies in SLE requires cognate interactions with autoreactive Th cells. This has been demonstrated as antibodies blocking MHCII abrogate autoantibody production in murine models 6. In this review we will discuss tolerance mechanisms of T cells and try to understand how the repertoire of Th cells affects autoimmunity. We will also discuss whether an autoreactive repertoire within Th cells causes a breakdown of immune tolerance or whether heightened activation of Th cells skews toward clonal expansion of self-reactive T cells.
Tolerance and repertoire selection during T development-central vs. peripheral
Development of mature immunocompetent T cells and the selection of their repertoire have been extensively reviewed 7. Therefore, the discussion of general T cell selection and development will be brief.
Selection in the thymus (central tolerance): It is accepted that the repertoire of naive T cells is determined through thymic selection. The recognition of peptides that are presented by thymic APCs is crucial for determining the fate of T cells; on one hand, it is essential for survival of thymocytes (positive selection). On the other hand, it leads to cell death for thymocytes (negative selection) or to skewing to regulatory T cell (Treg) differentiation. The affinity model of thymic selection emphasizes the strength of the interaction between the TCR and self-p-MHC complexes. Weak interactions are required for the protection of thymocytes from cell death by neglect and for promoting positive selection. Strong interactions, however, cause negative selection through apoptosis thereby preventing the maturation of self-reactive thymocytes (Figure 1). This model is supported by studies using TCR transgenic mice. Negatively selecting peptides are primarily high-affinity, whereas positively selecting peptides are primarily low-affinity 8. A surprisingly broad range of affinities are permissive for Treg differentiation 9.
Positive and negative selection occur in discrete areas: the cortex (positive selection) and the medulla (negative selection). Both compartments contain a microenvironment composed of different types of APCs, coordinating a spatial and temporal segregation of thymocyte selection. CD4/ CD8 double negative (DN) and double positive (DP) thymocytes undergo positive selection when they encounter peptide on cortical thymic epithelial cells (cTECs) 7. CTECs display a high rate of constitutive macroautophagy, which supports self-peptide loading to MHC II through an endogenous route 10, involving proteosomal activity, including PSMB11 11, cathepsin L 12 and thymus-specific serine protease (TSSP) 13.
Thymocytes with functional TCRs pass through positive selection and move to negative selection. A substantial portion of DP cells die through negative selection. Several types of APCs are involved in this process. Medullary TECs (mTECs) are a major player, but immune cells (dendritic cells and B cells) also play a significant role 14,15. mTECs express high levels of co-stimulatory molecules and peptides derived from non-thymic tissue antigens (named tissue-restricted antigens (TRAs)). The expression of TRAs is known to be regulated by at least two independent factors, Forebrain Embryonic Zinc Finger-Like 2 (FeZf2) and autoimmune regulator (Aire), which play complementary roles in TRA expression 16. MTECs can participate in negative selection directly by interacting with thymocytes or indirectly by transferring TRAs to neighboring immune cells, especially cDCs 17.
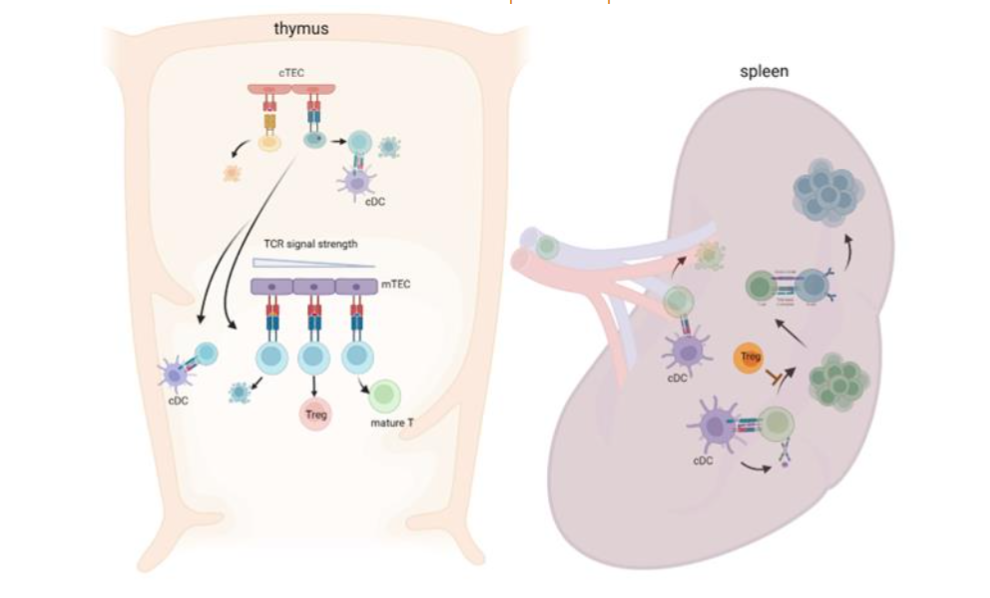
The role of different cell types in shaping negative selection and the TCR repertoire of thymocytes which emigrate from the thymus as naive T cells or Tregs is not fully understood.
Selection in the periphery: Thymic selection is a sophisticated process but imperfect and the pool of naïve T cells that exit the thymus contain both non-self-reactive cells that can react with foreign antigens (infections) and self-reactive T cells. Indeed, it has been estimated that as many as 20% of immunocompetent T cells are autoreactive 18. Therefore, removal or inactivation of autoreactive T cells continues in the periphery and signals from APCs and cues from the microenvironment are critical for this process. The strength of the TCR and p-MHC interaction, signals from co-stimulatory molecules and signals from soluble factors are all important in fate decisions of antigen-reactive T cells. It is not clear whether the composition of the naïve T cell repertoire is more critical for developing autoimmunity, or whether the repertoire of activated T cells in the periphery is more important.
The diversity of the naïve T cell repertoire does not necessarily positively correlate with the effectiveness of the immune response 19,20. A study analyzing antigen-specific TCR usage suggests that the efficiency of recruitment into the immune repertoire is related to TCR affinity for p-MHC and that a signaling threshold must be reached to ensure survival of recently activated naïve T cells 21,22.
In the periphery, DCs are the major APCs that activate naïve T cells and control T cell fate. Conventional DCs (cDCs) are potent APCs for both self-antigens and foreign antigens, participating in both immune tolerance and immunogenic responses (reviewed in 23). CDCs produce various pro- or anti-inflammatory cytokines, and these cytokines shape the immune response. The border between a tolerogenic and an immunogenic DC response is elusive, as shown by the cases in which tolerogenic DCs induce T cell activation rather than tolerance. For example, during colitis, CD103+ tolerogenic DCs can drive an inflammatory T cell response 24. The induction of an inflammatory T cell response is not observed in healthy mice following a transient treatment with toll like-receptor (TLR) agonist 25, suggesting that stimulation with transient TLR agonists alone are not sufficient to breach tolerance mechanisms and induce activation of autoreactive T cells.
Finding autoantigens and autoreactive T cells
Finding autoantigens and autoantigen-recognizing T cells is an important step to understand pathogenic mechanisms of autoimmunity. There is substantial evidence that colonization of commensal bacteria in the gut can trigger autoimmune disease, including EAE 26, RA 27,28, uveitis 29, and SLE 30. Gut microbes can affect the immune system locally and systematically by multiple mechanisms, including secretion of metabolites and effector T cell differentiation (reviewed in 31,32). Activation of self-reactive T cells can be triggered by exposure to a cross-reactive antigen derived from commensal microbiota. Pathogenic microbial antigen can also activate those T cells. It has been speculated that coxsackie virus triggers the activation of T cells cross-reactive with self-antigen in multiple sclerosis 33. T cells that have low affinity for self-antigen are not routinely activated by self-antigen. As memory T cells may respond to p-MHC with lower affinity than that needed to activate naive T cells, T cells may be activated by viral antigen and then reactivated as memory cells by self-antigen.
Research in SLE has focused on self-antigen recognizing T cells that are targeted by B cells. Identification of the nucleosome (histone/ DNA complex) as a primary antigen in SLE advanced our understanding of autoreactive T cells and the mechanisms that might drive pathogenic autoantibody responses in SLE. Early works identifying autoantigen-reactive helper cells (Th cells) were conducted by assessment of in vitro activation of autoreactive B cells. T cells and B cells were isolated from (NZBxNZW)F1 (NZF1) or (SWRxNZB)F1 (SNF1) lupus mouse models and co-cultured with peptides which were prepared from nucleosomes 34. T cells that induced autoantibody production from B cells were cloned. From these studies, critical peptide epitopes, which can breach tolerance, were mapped in H-2B (H2B10-33), H416-39 and H471-94. The epitopes are located at sites where the histones contact DNA in the nucleosome and are also targeted by B cells from lupus mice. Several lines of evidence suggest that the histone reactive Th cells may play a role in lupus development. Nucleosome-reactive Th cells arise spontaneously in lupus-prone mice, but not in normal mice. They appear many months before the development of a disease phenotype 35. The frequency of autoreactive Th cells is low in the pre-clinical stage but increases in diseased mice. Authors found that an immunization with nucleosome (a complex of DNA and histone), but not purified DNA or histone, can trigger the generation of pathogenic Th cells of SNF1 lupus mice in vivo, accelerating disease progression 35. Th cells exhibit a type 1 dominant response in immunized lupus mice, consistent with IgG2 and IgG3 being the isotypes of the induced autoantibodies 34. Histone peptide reactive T cells have also been found in SLE patients. Histone reactive Th clones were generated by immortalizing lupus CD4+ T cells with EBV and low-dose IL-2. Interestingly, there are overlapping epitopes in SLE patients and lupus mice. Moreover, distinct groups of histone peptides induce Th clones to secrete interferon (IFN)γ or IL-4/ IL-10 36. Similar studies have been performed with different nuclear antigens in lupus models and in SLE patients. Anti-Smith (Sm) antibodies are found in the sera of approximately 25% of individuals with SLE. The presence of anti-Sm antibodies is a very specific marker of SLE 37. Nine different polypeptides from the Sm molecule are antigenic and SmD polypeptides are regarded as the most antigenic within the Sm molecule to autoantibodies isolated from sera of MRL/lpr mice 38. Anti-SmD1 antibodies are present in NZF1 and MRL/lpr mice 39,40. T cells which recognize SmD1 peptide have been identified in both SLE patients and lupus-prone mice 41-43. Using an MHC II-peptide tetramer, U1-70 autoantigen reactive CD4+ T cells were found in MRL/lpr lupus mice 44. The frequency of U1-70 reactive CD4+ T cells positively correlate with disease progression. Further, these T cells produce IL-17 and IFNγ following ex vivo stimulation with peptides. Ribosomal P2 antigen reactive T cells were also characterized in SLE; the frequency of these T cells is significantly higher in anti-ribosomal P2 antibody-positive patients than anti-ribosomal P2 antibody-negative patients or healthy individuals 6. The ribosomal P2 antigen reactive T cells are CD4+ T cells and antigen-specific T cell proliferation is MHC II dependent. The presence of these antigen specific T cells in individuals with B cell reactivity to the same antigens suggests the T cells are Tfh cells.
The frequency of antigen-specific T cells within the human naïve repertoire has been estimated to range from 1 in 104 to 107 45. The limited number of known peptide epitopes and low frequency of antigen-specific T cells therefore permit only limited and potentially biased insight into the heterogenous T cell population reactive to autoantigens that may be present. The frequency and specificity of multiple autoantigen reactive CD4+ T cells were assessed using a T cell library and CD154+ enrichment to visualize autoreactive CD4+ T cells in ex vivo studies. Only patients with active SLE show an expansion of T cells with reactivity to multiple autoantigens; these T cells secrete IFNγ and IL-17 46. In the same study, autoreactive T cells were found in urine samples of patients with active lupus nephritis (LN), suggesting tissue infiltration of T cells which recognize self-antigens. The enrichment of antigen-activated T cells and the availability of epitope-searching algorithms has led to the discovery of additional autoantigens, vimentin and annexin 2, and the presence of vimentin/ annexin 2-recognizing Th cells was confirmed in blood and urine samples of patients with LN 47.
In summary, there are strong data that support the premise that autoreactive T cells exist in lupus mice and SLE patients (summary in (Table 1)). Those T cells are mostly CD4+ and are functionally competent since they induce B cells to produce autoantibodies and can modulate disease activity. They are heterogenous in terms of cytokine production, and their number positively correlates with disease activity.
| Antigen | epitope | Murine | SLE patients | references |
|---|---|---|---|---|
| Nucleosome | H471-94, H416-39, H2B 10-33, H391-105 | SNF1, BWF1 | active SLE | 48, 36 |
| Sm/ Sm U1RNP | U1-70131-150 | MRL/lpr | SLE (MCTD) | 44 |
| Ro | active SLE | 46 | ||
| La | active SLE | 46 | ||
| HMG | SLE | 49 | ||
| hnRNP2 | SLE | 50, 6 | ||
| vimentin | Lupus nephritis | 47 |
Treg repertoire development and autoimmunity
The generation of Tregs is required for maintaining immune tolerance and FOXP3- CD4+ T cells. When patients lack Tregs due to a FOXP3 deficiency, they develop immune dysregulation polyendocrinopathy, enteropathy X-linked (IPEX) syndrome 51. The generation of Tregs occurs both in thymus and in the periphery, and both kinds of Tregs are required for the maintenance of immune homeostasis 52,53. The development of Tregs depends on a TCR interaction of high affinity for the cognate antigen 54. Observations made from Nur77-EGFP reporter mice, where the relative level of TCR signaling could be inferred from the level of EGFP expression, indicated that Tregs undergo stronger TCR signaling than non-Tregs during thymic development 55. Tregs seem to require specificities distinct from those of non-Tregs for their differentiation 54,56. Cognate antigen dependent development of self-antigen reactive Tregs was demonstrated in myelin oligodendrocyte glycoprotein (MOG)-deficient mice. When the MOG-deficient mice were reconstituted with hematopoietic stem cells expressing a MOG-reactive TCR, MOG-reactive Treg development was severely reduced but MOG-reactive conventional T cell development was not affected 57. Thus, autoreactive Tregs are positively selected on endogenous self-antigen.
It is accepted that the reduced number of Tregs and reduced suppressive function of Tregs can facilitate the development of autoimmune diseases including SLE, however whether the repertoire of Tregs affects immune homeostasis is understudied. The repertoire of Tregs is diverse and the mechanisms of generation of Tregs with diverse TCRs is partly driven by FOXP3 expression in Treg precursors. FOXP3 expression is a key determinant for Treg differentiation, and the intronic Foxp3 enhancer conserved noncoding sequence 3 (CNS3) is required for increasing the TCR diversity of Tregs. CNS3 works as an epigenetic switch that confers a poised status to the Foxp3 promoter to make Treg lineage commitment responsive to a broad range of TCR stimuli. Deletion of CNS3 in Tregs increases TCR signal strength and proliferation but decreases diversity of the repertoire, leading to a development of autoimmunity 58. Importantly, no difference is seen in the repertoire in non-Tregs, demonstrating the importance of antigen specificity of Treg in maintaining immune homeostasis.
TCR-antigen interactions of different strengths can lead to the development of distinct Treg subsets, including GITRhiPD1hiCD25hi (Triplehi Treg) and CITRloPD1loCD25lo (Triplelo Tregs). These Treg subsets differ by their expression of Helios, ICOS, CD44 and CD62L 59, and affinity for self-antigen: Triplehi Tregs express TCR of higher affinity for self-antigen than Triplelo Treg. They also display different functions in vivo, Triplehi Tregs control T and B cell activation in secondary lymphoid organs, while Triplelo Tregs protect mice from development of colitis 59. A recent study suggested autoreactive Tregs as therapeutics in SLE 60. In this study, authors identified HLA-DR15 restricted immunodominant epitopes from the Sm antigen, generating Sm-specific Tregs. These engineered Sm specific Tregs suppressed a proinflammatory cytokine producing effector T cells and a disease progression in a humanized mouse model of LN. This study underscores importance of peripheral tolerance mechanism by self-antigen reactive Tregs.
TCR repertoire in lupus mouse models
Repertoire selection is inevitably related to the immune fitness of the host. The selection process throughout T cell development (including both positive and negative selection in thymus and in peripheral lymphoid organs) can affect the repertoire of T cells and antigen-specific responses in the periphery. Lack of cathepsin L or TSSP in cTECs during positive selection results in aberrant CD4+ T cell selection; cathepsin L-deficient mice have decreased polyclonality of the CD4+ T cell repertoire 12, whereas TSSP-deficient mice have normal clonality but show defective positive selection of certain MHC II-restricted TCRs and altered antigen-specific CD4+ T cell responses in the periphery 13. A direct role of TCR repertoire in autoimmunity has been seen with altered negative selection. Mutations in AIRE cause autoimmune polyendocrinopathy-candidasis-ectodermal dystrophy (APECED), a severe multi-organ autoimmune disease 61. Aire-deficient mice have an expanded TCR repertoire and develop the features of APECED phenotype 62. Monoallelic variants in NFKB2 that lead to loss of function can result in immunodeficiency and many patients with these variants display at least one autoimmune manifestation 63. Since NFkB2 is required for AIRE expression and for the development of mTECs 64,65, the breakdown of tolerance in NFkB2 deficiency results in part from altered central tolerance. There is also evidence that perturbed tolerance can be due to altered TCR signaling. Mutations in the TCR signaling pathway, such as loss-of function mutations in the linker for activation of T cells (LAT), lymphocyte-specific protein tyrosine kinase (LCK) deficiency, or autosomal-recessive CD3g deficiency can result in autoimmunity 66-68. Mutations in all 3 genes lead to severe T lymphopenia but direct evidence of an altered repertoire with negative selection of self-antigen reactive T cells has not been demonstrated except in mice with the LAT mutation. Mice with a LAT mutation (Y136F), which precludes docking of PLCg on LAT, develop lupus-like autoimmunity. LAT mutant mice with a transgenic TCR show an enhanced TCR-mediated activation and altered thymocyte selection, resulting in release of HY-reactive T cells, which would normally be eliminated through negative selection 69. We observed that alterations in peripheral APCs can distort the T cell activation to promote lupus-like autoimmunity in mice with a DC-specific deletion of B lymphocyte induced maturation protein (BLIMP)-1 without affecting the TCR repertoire of naive T cells 70.
Evidence of defective central tolerance or a disrupted T cell repertoire in many lupus prone mice models has been suggested but not fully understood. Early studies focused on characterizing the repertoire of T cells which react with known autoantigens and identified groups of V genes of TCRs that are expanded in autoreactive T cells in SNF1 lupus mice. CD4+ T cells which can induce autoantibody production from B cells showed an increased frequency of Vβ8 or Vβ4 71. In the DWEYS-peptide (peptide cross-reactive with dsDNA) induced lupus mouse model, TCR V gene usage differed between autoantigen reactive CD4+ T cells and total CD4+ T cells, with Vβ8 dominance followed by Vβ2/4/6/10 in total CD4+ T cells, but a Vβ6 dominance in DWEYS-specific T cells 72. Usage of the TCR Vβ8 increases over time in MRL/lpr mice 73. The autoantigen for Vβ8 is not identified, but the usage of Vβ elements reflects the stage of the disease which might suggest a clonal selection of autoantigen specific TCR in the lupus mice. Moreover, skewed V gene usage and CDR3 sequences were found in tissue infiltrating CD4+ T cell in MRL/lpr mice. Moore and colleagues compared the clonality in CD4+ T cells in brain choroid plexus in MRL/lpr mice compared to MRL/MpJ control mice and to spleen and salivary gland from MRL/lpr mice. They found a significant difference in the TCR repertoire between MRL/lpr and MRL/MpJ, but also among MRL/lpr tissues. The TCR repertoire in choroid plexus exhibited greater clonality and sequence homology compared to spleen and salivary gland 74. Kidney-infiltrating CD4+ T cell clones from MRL/lpr mice have high levels of CD5 and TCR and respond to splenic DCs but not B cells or macrophages in MHC II-restricted manner. When those T cell clones are adoptively transferred, they promoted lupus development and nephritis suggesting that they are autoreactive 75.
NZF1 mice develop T cell dependent autoantibody production spontaneously, but T cells exhibit normal central tolerance without a gross alteration in repertoire 76. Autoantibody production rather depends on abnormal activation of T cells in the periphery. Hyperactivation of T cells in NZF1 mice results in part from defects in Treg function 77. Our group found that the diversity of follicular helper T (Tfh) cells is increased in lupus-prone compared to control mice in a model of spontaneous lupus secondary to BLIMP1 deletion in cDCs 78. The increased repertoire of Tfh cells and development of a lupus phenotype is associated with increased activity of cathepsin S in cDCs. Thymic selection and the repertoire of SP CD4 thymocytes are not affected in this model, suggesting intact central tolerance. The repertoire of naive CD4+ T cells in the periphery is similar to control mice, but stimulation with peptide loaded BLIMP1-deficient cDCs induces a stronger TCR stimulation with an increased TCR diversity within the activated naive T cells 70. These data underscore the important role of antigen presentation by cDCs in the selection of activated T cells in the periphery and the contribution of aberrantly enhanced T cell activation to autoimmunity. Studies of another B cell driven lupus mouse model (mixed bone marrow chimera of 564Igi mice which an autoreactive B cell receptor (BCR)) showed that Tfh cells from the autoimmune donor are distinct from non-autoimmune Tfh cells in transcriptional profile and repertoire 79. Tfh cells from autoimmune mice express many genes involved in activation and exhaustion. Paired TCR repertoire of Tfh cells revealed that CDR3 length, V gene usage or clonal expansion are not different, but repertoire of lupus Tfh cells is distinguished from that of non-lupus mice.
TCR repertoire in individuals with SLE
Although the TCR repertoire in an individual is highly diverse 80,81, autoimmune conditions can cause clonal expansion of certain T cells. The widespread use of high throughput next-generation sequencing (NGS), especially RNA-seq, significantly advances our information on repertoire and T cell activation state. Results obtained from high throughput NGS, however, are somewhat inconsistent among patient studies at present. A longitudinal analysis of blood T cell repertoire of individuals with SLE (n=11) showed increased clonality in individuals with SLE compared to healthy individuals, and the clonality was not altered by disease activity 82. Unlike earlier studies from animal models, there was no clear preference in Vβ or Jβ gene usage among the top 100 expanded clones from individuals with SLE compared to healthy individuals. Another study identified clones associated with disease severity in a large number of patients and showed more clonal expansion in autoimmune patients; in contrast another study compared TCR repertoire from blood T cells of patients with SLE (n=877), patients with rheumatoid arthritis (RA) (n=206) and healthy individuals (n=439) and demonstrated a significant difference in Vβ gene usage between SLE or RA and healthy individuals. This study also compared the repertoire between SLE and RA and found a high similarity within the disease-associated clones. These disease-associated clones were expanded with disease severity 83. Hou and colleagues analyzed the TCRb and BCR heavy (H)-chain repertoire of blood cells from patients with individuals with SLE (n=10) compared to healthy individuals (n=10) 84. They found a significant reduction in diversity of BCR-H and BCR-H CDR3 length in SLE patients, but not in TCR repertoire in individuals with SLE, including diversity and CDR3 length. Instead, they found skewed V gene usage and CDR3 sequence in both TCRβ with more TRBV6-5, TRBV11-3, TRBV6-3, TRBV6-1, TRBV27 and TRV28 and BCR-H with more IGHV3-49, IGHV2-70 and IGHV2-5 in SLE patients. The inconsistencies among these studies need to be resolved. The studies were conducted with total blood T and B cells, and cell subsets were not considered. More recent studies have analyzed immune cells in a more comprehensive way by using scRNA-seq, with a combined analysis of gene expression and immune repertoire at a single cell level. Wu and colleagues compared TCR and BCR in 2 different ethnicity groups of patients with SLE and healthy controls. They confirmed the increased TCR and BCR clone numbers in patients with SLE compared to healthy controls, but diversity was significantly decreased for both TCR and BCR of patients with SLE. The authors also attempted to find TCR repertoire changes within subsets, but none were identified, perhaps due to a limited sample size 85.
Conclusion
Developing a diverse repertoire of T cells is important for protective immunity, but self-reactive T cells endanger the host and contribute to autoimmune diseases. It is well accepted that all the processes involved in T cell selection, including positive selection and negative selection and Treg differentiation in the thymus and further selection in the periphery, are critical for controlling self-reactivity. In addition to the repertoire of T cells, activation status of T cells can affect the fate of autoantigen-recognizing B cells in the periphery. Activated T cells or self-reactive T cells can secrete pro-inflammatory cytokines, leading to immunoglobulin secretion with pathogenic, class-switched isotypes. So far, most data about TCR repertoire from patients with SLE or lupus mouse models have been generated from total CD4+ T cells and repertoire of effector T cells (especially Tfh cells) or Treg are not well described. How the repertoire of T cells is affected by antigen challenge and whether this process differs between patients with SLE, and healthy individuals is also not known.
Conflict of interest statement
The authors have declared no conflicts of interest to declare.
Funding statement
This work was supported by the NIAMS, US National Institute of Health R01 AR074565 to SJK and BD.
Acknowledgement
We are grateful to George Georgiou at the University of Texas at Austin for constructive discussion for the preparation of manuscript.
References
- Roth DB. V(D)J Recombination: Mechanism, Errors, and Fidelity. Microbiol Spectr. Dec 2014;2(6)doi:10.1128/microbiolspec.MDNA3-0041-2014
- Rudolph MG, Stanfield RL, Wilson IA. How TCRs bind MHCs, peptides, and coreceptors. Annu Rev Immunol. 2006;24:419-66. doi:10.1146/annurev.immunol.23.021704.115658
- Suarez-Fueyo A, Bradley SJ, Tsokos GC. T cells in Systemic Lupus Erythematosus. Curr Opin Immunol. Dec 2016;43:32-38. doi:10.1016/j.coi.2016.09.001
- Li H, Boulougoura A, Endo Y, Tsokos GC. Abnormalities of T cells in systemic lupus erythematosus: new insights in pathogenesis and therapeutic strategies. J Autoimmun. Oct 2022;132:102870. doi:10.1016/j.jaut.2022.102870
- Malkiel S, Jeganathan V, Wolfson S, et al. Checkpoints for Autoreactive B Cells in the Peripheral Blood of Lupus Patients Assessed by Flow Cytometry. Arthritis Rheumatol. Sep 2016;68(9):2210-20. doi:10.1002/art.39710
- Crow MK, DelGiudice-Asch G, Zehetbauer JB, et al. Autoantigen-specific T cell proliferation induced by the ribosomal P2 protein in patients with systemic lupus erythematosus. J Clin Invest. Jul 1994;94(1):345-52. doi:10.1172/JCI117328
- Klein L, Hinterberger M, Wirnsberger G, Kyewski B. Antigen presentation in the thymus for positive selection and central tolerance induction. Nat Rev Immunol. Dec 2009;9(12):833-44. doi:10.1038/nri2669
- Kyewski B, Klein L. A central role for central tolerance. Annu Rev Immunol. 2006;24:571-606. doi:10.1146/annurev.immunol.23.021704.115601
- Lee HM, Bautista JL, Scott-Browne J, Mohan JF, Hsieh CS. A broad range of self-reactivity drives thymic regulatory T cell selection to limit responses to self. Immunity. Sep 21 2012;37(3):475-86. doi:10.1016/j.immuni.2012.07.009
- Nedjic J, Aichinger M, Emmerich J, Mizushima N, Klein L. Autophagy in thymic epithelium shapes the T-cell repertoire and is essential for tolerance. Nature. Sep 18 2008;455(7211):396-400. doi:10.1038/nature07208
- Florea BI, Verdoes M, Li N, et al. Activity-based profiling reveals reactivity of the murine thymoproteasome-specific subunit beta5t. Chem Biol. Aug 27 2010;17(8):795-801. doi:10.1016/j.chembiol.2010.05.027
- Nakagawa T, Roth W, Wong P, et al. Cathepsin L: critical role in Ii degradation and CD4 T cell selection in the thymus. Science. Apr 17 1998;280(5362):450-3. doi:10.1126/science.280.5362.450
- Gommeaux J, Gregoire C, Nguessan P, et al. Thymus-specific serine protease regulates positive selection of a subset of CD4+ thymocytes. Eur J Immunol. Apr 2009;39(4):956-64. doi:10.1002/eji.200839175
- McCaughtry TM, Baldwin TA, Wilken MS, Hogquist KA. Clonal deletion of thymocytes can occur in the cortex with no involvement of the medulla. J Exp Med. Oct 27 2008;205(11):2575-84. doi:10.1084/jem.20080866
- Melichar HJ, Ross JO, Herzmark P, Hogquist KA, Robey EA. Distinct temporal patterns of T cell receptor signaling during positive versus negative selection in situ. Sci Signal. Oct 15 2013;6(297):ra92. doi:10.1126/scisignal.2004400
- Takaba H, Morishita Y, Tomofuji Y, et al. Fezf2 Orchestrates a Thymic Program of Self-Antigen Expression for Immune Tolerance. Cell. Nov 5 2015;163(4):975-87. doi:10.1016/j.cell.2015.10.013
- Hubert FX, Kinkel SA, Davey GM, et al. Aire regulates the transfer of antigen from mTECs to dendritic cells for induction of thymic tolerance. Blood. Sep 1 2011;118(9):2462-72. doi:10.1182/blood-2010-06-286393
- Yu W, Jiang N, Ebert PJ, et al. Clonal Deletion Prunes but Does Not Eliminate Self-Specific alphabeta CD8(+) T Lymphocytes. Immunity. May 19 2015;42(5):929-41. doi:10.1016/j.immuni.2015.05.001
- La Gruta NL, Rothwell WT, Cukalac T, et al. Primary CTL response magnitude in mice is determined by the extent of naive T cell recruitment and subsequent clonal expansion. J Clin Invest. Jun 2010;120(6):1885-94. doi:10.1172/JCI41538
- La Gruta NL, Thomas PG, Webb AI, et al. Epitope-specific TCRbeta repertoire diversity imparts no functional advantage on the CD8+ T cell response to cognate viral peptides. Proc Natl Acad Sci U S A. Feb 12 2008;105(6):2034-9. doi:10.1073/pnas.0711682102
- Gascoigne NR, Rybakin V, Acuto O, Brzostek J. TCR Signal Strength and T Cell Development. Annu Rev Cell Dev Biol. Oct 6 2016;32:327-348. doi:10.1146/annurev-cellbio-111315-125324
- Zikherman J, Au-Yeung B. The role of T cell receptor signaling thresholds in guiding T cell fate decisions. Curr Opin Immunol. Apr 2015;33:43-8. doi:10.1016/j.coi.2015.01.012
- Fucikova J, Palova-Jelinkova L, Bartunkova J, Spisek R. Induction of Tolerance and Immunity by Dendritic Cells: Mechanisms and Clinical Applications. Front Immunol. 2019;10:2393. doi:10.3389/fimmu.2019.02393