





Calpain Functions in Disease: Insights for Medical Practice
Plant and animal calpain functions, association with microtubules and possible medical applications
Hilde-Gunn Opsahl-Sorteberg¹*, Espen Evju¹, Zhe Liang² and Jennifer C. Fletcher³
- Norwegian University of Life Sciences, Biosciences, Ås, Norway
- Chinese Academy of Agricultural Sciences, Beijing, China
- Plant Gene Expression Centre, USDA/UC Berkeley, Albany CA, USA
OPEN ACCESS
PUBLISHED:30 September 2024
CITATION: Opsahl-Sorteberg, H., Evju, E., et al., 2024.
Plant and animal calpain functions, association with microtubules and possible medical applications. Medical Research Archives, [online] 12(9).
https://doi.org/10.18103/mra.v12i9.5639
COPYRIGHT: © 2024 European Society of Medicine. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
DOI https://doi.org/10.18103/mra.v12i9.5639
ISSN 2375-1924
Abstract
Calpains are calcium-activated cysteine proteases that activate a vast variety of substrates by cleavage. First reported in 1964, calpains are found in both prokaryotes and eukaryotes and display disparate multidomain architectures. The term calpainopathy was coined in 1995 when calpains were first linked to cell cycle control and cancer, and since then calpains have been implicated in many additional medical conditions including heart disease, multiple sclerosis, diabetes, sickle cell disease, and various neurological disorders. The evolution of calpains is under active investigation, but the core CysPc cysteine protease domain can be traced back to bacteria and the membrane associated MIT domain to Archaea. Here we review calpain evolution and suggest that the MIT-CysPc domain was present in the first eukaryotic common ancestor, and that this diverged into a minimum of four independent last eukaryotic common ancestors making up diverse groups from animals to land and marine plants. How calpains function at the cellular level is likewise not fully resolved. However, they are recognized to play roles in cell division, adhesion, fusion, proliferation, migration and signaling in animals and to act on stem cell functions via microtubules in land plants. Just recently calpains have also been connected to the microtubule organizing center in land plants and brown algae. We present a possible basic function of calpain domains, their connections to membranes and a possible calcium channel, supported by an updated phylogeny. Finally, we provide an overview of human calpains, potential functions and medical conditions to which they are linked and suggest possible development of calpain transcriptomic diagnostics to increase medical precision and treatment. We believe that understanding calpains has promising medical spinoffs and look forward to seeing this field unfold in the years to come.
Keywords
Calpain, calcium, cell division, microtubule, disease, cancer
Introduction
Calpains are calcium-activated cysteine proteases that alter the functions of their numerous substrates by cleavage. The name calpain combines ‘cal’ and ‘pain’, which are derived from calcium and cysteine proteases such as papain and legmain. The calpain system is one of four cellular proteolytic systems along with the proteasome, lysosome and caspase systems and is involved in many cellular processes in multicellular eukaryotes. The 15 human calpains are involved in many of our most common and serious diseases such as cancer and stem cell functions by regulating the cell cycle, neural functions linked to dementia, and oxygen transport linked to sickle cell disease. Most research on calpains has been done on animals and within medical science, yet our understanding of their biological functions is still only rudimentary.
Members of the calpain family are multi-domain proteins that share a conserved core cysteine protease (CysPc) catalytic domain as well as diverse combinations of additional domains. Mammalian calpains are divided into classical and non-classical calpains based on their domain organization. The nine classical calpains, represented by calpain-1 and calpain-2, are composed of a larger and a smaller subunit (Table 1). The larger subunit is comprised of an N-terminal anchor helix, a CysPc domain, a calpain-type beta-sandwich (CBSW) domain and a penta-EF-hand (PEF) domain. The smaller subunit consists of only a PEF domain and a glycine-rich (GR) hydrophobic region. Following activation by calcium ions, these subunits assemble to form a functional heterodimer with catalytic activity. The six non-classical calpains consist of a single large subunit and typically function as monomers. These calpains lack the PEF domain and in some cases the CBSW domain. Other domains found in various mammalian calpains include a microtubule interacting and transport motif (MIT), a C2 or C2-like (C2) domain, and a zinc-finger (Zn) motif.
The calpain gene superfamily is evolutionarily ancient and its members are widely distributed among both prokaryotic and eukaryotic lineages.
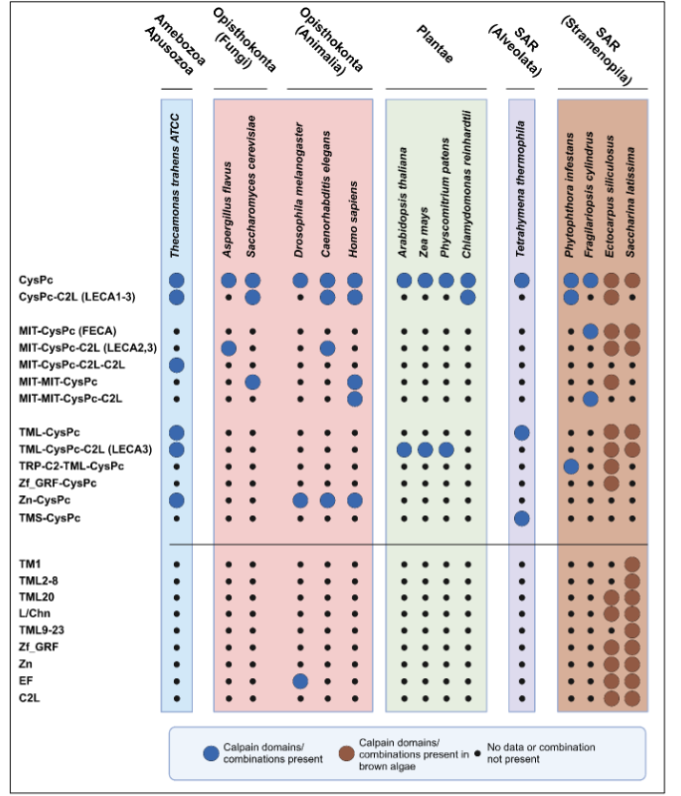
Sequences encoding the core CysPc domain are present in eubacteria, including cyanobacteria, but have not been identified in Archaea. Genes encoding multiple calpain family members have also been cataloged in fungi, invertebrates, and mammals, in unicellular eukaryotes such as Thecamonas trahens and Tetrahymena thermophila, and in several macroalgal species. In contrast, land plants have one unique calpain gene, named DEFECTIVE KERNEL1 (DEK1). Interestingly, the genome of the brown macroalga Saccharina latissima contains sequences similar both to the land plant DEK1 gene as well as to many of the calpain versions found in animals.
| Calpain Gene Number | Suggested Functions |
|---|---|
| CAPN 1&2: Classical | Expressed: Ubiquitously. Membrane fusion, platelet activation, cell cycle progression, cytoskeletal remodeling, cleavage of receptors, neurons functions, cell adhesion, retinal apoptosis. Calpain-1 protects while -2 is neurodegenerative, where they balance each other’s brain function affecting memory. |
| CAPN 2: Classical | Age added effects of UV-damage, high sugar levels if diabetic, environmental toxins and more. Can compromise membrane proteins possibly through increased ion permeability in lens fiber cells. Embryo development. |
| CAPN 3: Classical | Expressed: Skeletal muscle. Expressed in skeletal muscle, muscular dystrophy, nuclear localization. Role in Ca2+ release independent of its protease activity. Forms homodimers and trimers. |
| CAPN 5 & 6: Non-classical | Expressed: Ubiquitously. Homologue of C. elegans sex determining gene TRA-3, exp most tissues, but especially in the central nervous system. Has 3 extended loops possibly explaining the need for higher Ca2+ levels for activation. |
| CAPN 7: Non-classical | Expressed: Ubiquitously. Divergent sequence, more related to the fungal calpain Aspergillus nidulans. |
| CAPN 8&9: Classical | Expressed: Gastrointestinal tract. Form heterodimers, CAPN8 can also form homodimers, SNPs known to inactivate them can be used as diagnostics. CAPN9 forms heterodimer with CAPN1. G1 cell cycle arrest and caspase-mediated apoptosis, tumor suppressing role by degradation tract-specific oncogenes. |
| CAPN 10: Non-classical | Expressed: Ubiquitously. Insulin-mediated glucose turnover, cellular apoptosis, renal cell viability, tubule repair in renal cells. Regulator of glucose metabolism, thereby associated with development of type 2 diabetes. Requires special intracellular localization or interacting partner(s) to acquire proteolytic activity and cleaved by calpain-2. |
| CAPN 11: Classical | Expressed: Testis. Expressed in testis from 14 days after birth during pachytene spermatocyte development, suggested function during meiosis. |
| CAPN12-14: Classical | Expressed: Hair follicle cells, ubiq and gastrointestinal. Unknown functions. |
| CAPN 15 & 16: Non-classical | Expressed: Ubiquitously, unknown functions. |
The aim of this article is to review the evolutionary origin of calpains, their biological functions in land plants and animals, and their association with human medical conditions and diseases. We present an overview of recent progress in understanding calpain activity from previous studies in animals and land plants, as well as new discoveries in macroalgae that suggest biological similarities to both animal and plant calpains at the evolutionary and functional levels. Based on these comparisons we find increased support for a core calpain function in orchestrating microtubule orientation within cells. Finally we discuss the roles of calpains in human cell biology and disease, and suggest that consideration of calpain functions in diverse species can add valuable understanding to improve medical applications of calpain-based therapies.
Evolutionary origin of calpains
Four ancestral calpains date more than a billion years back. Extensive phylogenetic studies of calpains seeking their evolutionary origin and possible functional implications suggest the cysteine protease domain originates from bacteria, and further that the large variation in domain architecture between and within eukaryotic species descends from four original versions early in eukaryotic history. These were likely present before the split of the eukaryotic supergroups, the 1) Apusonomidae, containing Thecamonas, 2) Opistokonta, containing fungi and animals, 3) SAR, containing Tetrahymena and the brown algae Ectocarpus and Saccharina, and 4) Plantae, containing land plants. The unicellular eukaryotes from which these different supergroups evolved underwent a massive expansion of the four original calpains into 41 different domain combinations. This extensive structural variation most likely arose from a combination of duplications, domain shuffling and secondary domain loss or modification during the course of evolutionary selection.
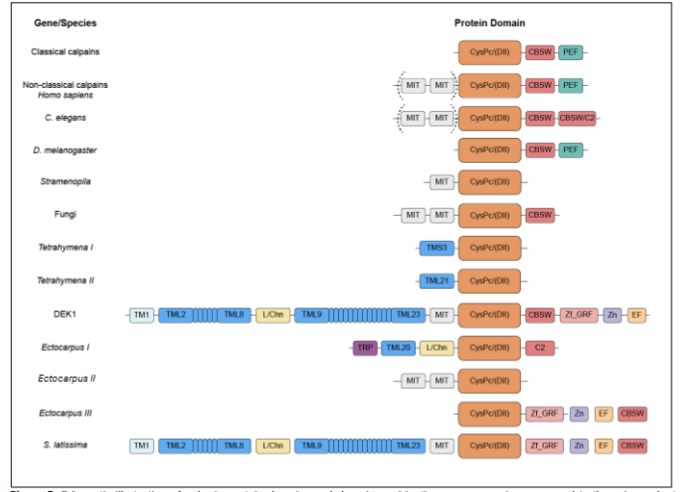
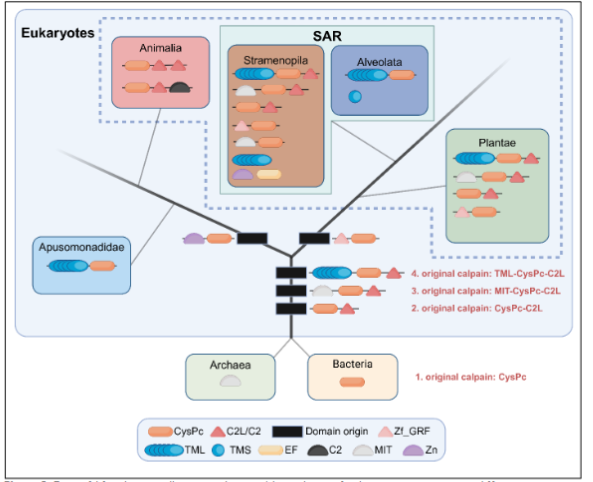
The membrane anchoring activity of the DEK1 calpain might have been retained in land plants due to their cell walls, a physical barrier not found in animals, possibly meaning linking directly to the membranes is not as important in organisms without cell walls. It is still striking how Thecamonas and metazoans have MIT suggested functioning on the cytoskeleton and possibly also connecting the cytosolic calpains to membranes, while the Tetrahymena and land plant SAR lack MIT. This might possibly explain why TML anchoring to the membranes is important in land plants, a function replacing the MIT domain in other organisms. Both are consistent with central functions by the cytoskeleton for calpain function via the MIT or TML. Thecamonas and brown algae are so far the only eukaryotes found to have retained both the MIT and TML domains, possibly allowing some branches to have kept the MIT while plants kept the TML. The TML anchors the calpain to the cell membranes, such as the outer cell membrane and nuclear envelope. Calpain membrane activity has only been studied in land plants and might give us insight into the possible basic function of calpains, including those in animals.
We suggest there existed three phylogenetically distinct membrane-bound calpains in the LECAs, now present in land plants, brown algae and animals. The membrane-associated land plant calpain, DEK1, is activated by Ca2+ in the presence of an experimentally added channel. The substantial membrane part of DEK1 has also been suggested to possibly contain a Ca2+ channel region within it, potentially explaining why the membrane anchor has been retained in this calpain through evolution. DEK1 is indicated to be functionally involved in regulating the internal, cytosolic portion of its calpain through a Ca2+ channel further linked to the organization of MT. The brown macroalga Ectocarpus and Saccharina both contain sequences similar to the transmembrane domains of the land plant DEK1 calpain as well as to many of the calpain versions found in animals, suggesting that macroalgal calpain proteins may be composites in which both animal- and plant-associated domains were retained.
In animals the cytosolic calpains, although not bound to membranes, are consistently reported to be associated with them. The animal calpain-1 and calpain-2 proteins are bound to Transient Receptor Potential (TRP) channels and even activate TRPC5. The TRP channels make up a superfamily of Ca2+ permeable nonselective membrane cation channels and contribute to intracellular Ca2+ fluxes and cell signaling. These channels often form units of 24 transmembrane pores specific to eukaryotes. TRP channels have not been identified in land plants but are present in the closely related green algae, opposite to DEK1, suggesting that TRP channel activity might replace DEK1 function in green algae.
Functions of plant calpains to set 3D cell orientation via microtubules
The unique land plant calpain gene DEK1 was first identified from a maize (Zea mays) mutant collection having aborted seeds, shown to be due to loss of epidermal cells and embryo arrest at the globular stage. We further found the gene to be expressed in most cells, with higher expression in actively dividing cells and in stem cells in particular. Arabidopsis dek1 mutant embryos have defective cell division patterns and planes, and in more severely affected embryos, nearly all the cell walls are incorrectly positioned. These defects are associated with disorganized arrangements of microtubules (MTs) within the cells, demonstrating that DEK1 regulates the arrangement of the cortical MT systems during early embryo development, which in turn may affect cell wall deposition. Together the data suggest that DEK1 sets the 3D cell division orientation for plant embryos to progress beyond globular stage by positioning cell walls according to their microtubule orientation.
DEK1 also affects the cell divisions in the developing suspensor. In land plants the upper tier of the dividing zygote develops into the embryo proper, while the lower tier forms an extra-embryonic suspensor structure connecting it to the surrounding endosperm tissues. The suspensor divides exclusively through anticlinal cell divisions before dying by programmed cell death, whereas the embryo proper divides in various planes to set the 3D orientation developing the new plant generation. In Arabidopsis dek1 mutants, the suspensor develops like the embryo proper by dividing in the periclinal orientation and expressing genes otherwise only present in the apical embryo. This suggest repressive signals from the embryo proper are needed to keep the suspensor from carrying out its embryogenic potential. One such marker is WOX2, the expression of which in the apical cells depends on a first asymmetric cell division of the zygote. dek1 mutants fail to express WOX2, likely due to a failure of the microtubule-orientated asymmetric first cell division. In addition, expression of the hormone transport protein PIN4, which is normally restricted to the suspensor, occurs ectopically in the dek1 embryo proper.
The lack of proper PIN4 protein localization might be a factor in why dek1 mutants are unable to repress proper embryo development. Medical science can achieve important insights from comparing the functions of evolutionary-related genes in different species. DEK1 is an ideal candidate to study calpain activity, because it occurs as a single copy in plants whereas multigene calpain families and the many varying sequences in other important eukaryotes complicate functional studies. To date the plant studies are consistent with a fundamental role for DEK1 in directing asymmetric cell divisions, in particular when shifting from 2D to 3D orientations, and in activating the expression of genes that depend on asymmetric divisions for their spatial patterning. The requirement of DEK1 for correct orientation of cell divisions is associated with a function in regulating MT organization, indicating a core role for DEK1 in cytoskeletal control. We suggest DEK1 acts by organizing the MT and argue this could be a core function of calpains in animals also. Animals have both centrosomes and microtubule organizing centres (MTOC), as do brown algae, whereas land plants lack centrosomes and therefore depend on the MTOC to organize the cytoskeleton throughout the cell cycle and to control cell division.
Calpain association with medical conditions – utility in diagnostics and therapy
In animals calpains have been linked to a wide spectrum of functions including calcium regulation, signal transduction, cytoskeleton dynamics, cell mobility, cell cycle progression, long-term potentiation in neurons, muscle protein break-down, cell adhesion, cell fusion and apoptosis. Calpains are known to be involved in pathophysiological mechanisms and specifically in human medical conditions such as cancer, Alzheimer’s disease, multiple sclerosis, Parkinson’s disease, Huntington’s disease, Type2 diabetes, estrogen-mediated cancer metastasis, aging related syndromes, sickle cell disease, asthma, cardiac dysfunction and blindness, and even alcoholism and malaria. These pathological functions are associated with either reduced or elevated calpain activity, leading to decreased or excessive substrate cleavage, respectively.
The classical calpains are numbered calpain-1, 2, 3, 8, 9, 11, 12, 13 and 14. Calpain-1 and calpain-2 function in membrane fusion, platelet activation and cell cycle progression, cytoskeletal remodeling, and cleavage of receptors such as epidermal growth factor (EGF) in neurons and the eye especially. Calpains have been shown to reduce function in retinal apoptosis, by downregulating proapoptotic proteins and NF-kappaB and thereby neuroprotecting retinal ganglion cells. Together with caspase-3 they can cleave cytoskeletal, cytosolic and nuclear substrates. Calpain-1, calpain-2 and caspase-3 further have synergistic effects on acute neuronal cell death, while surprisingly Calpain 1 and 2 have opposite effects. Calpain-1 is neuroprotective while calpain 2 is degenerative, suggesting downregulation of calpain-2 may be a target for neural degenerations including traumatic brain injury and repeated concussions. Calpain inhibition has further been shown to slow down cataract formation by protecting cytoskeletal proteins from calpain proteolysis.
Calpain-1 and -2 are linked to neurological disorders – stroke and Alzheimer’s disease (AD) – as well as several types of cancer. They are found to be upregulated in breast cancer cells and to suppress cancer treatment effects, but this differs between cancer types and calpains. Knockout of CAPN1 also leads to neurological disorder spastic paraplegia. In addition, Calpain-1 and -2 have been shown to cause disassociation of tau from MT, thereby leading to neuronal death, making them targets for treatment of Alzheimer’s disease. Calpain-2 is reported to cause added effects of UV-damage, high sugar levels if diabetic, environmental toxins and to compromise membrane proteins possibly through increased ion permeability in lens fiber cells. In mice, calpain-2 is also needed for embryo development.
| Calpain Number | Medical Association |
|---|---|
| Calpain-1 | Neurological disorders such as stroke, Alzheimer’s disease (AD), spastic paraplegia. |
| Calpain-2 | Transcript marker of sudden cardiac death and a potential therapeutic target in various forms of neurodegeneration, including traumatic brain injury and repeated concussions. |
| Calpain-1&2 | Upregulated in breast cancer cells and suppresses cancer treatment effects. Cause disassociation of tau from MT – neuronal death therefore targets for AD treatment. Calpain inhibition has been shown to slow down cataract formation in humans. |
| Calpain-3 | Limb-girdle muscular dystrophy (LGMD). Highly expressed in melanoma cell lines and bovine bladder tumors. |
| Calpain-5 | ADNIV. |
| Calpain-9 | Gastric cancer, suppression of tumorigenesis. |
| Calpain-10 | Type 2 diabetes (T2D). |
| Calpain-11 | Functions during meiosis for sperm function. |
| Calpain-12 | Developmental eye disorders caused by disruption of the optic fissure disclosure. |
| Calpain-14 | Eosinophilic esophagitis. |
CAPN8 and CAPN9 are expressed in the gastrointestinal (GI) tract, can be inactivated by specific SNPs, form dimers with each other and can be used as diagnostics for gastric cancer. Calpain-9 is linked to G1 cell cycle arrest and caspase-mediated apoptosis and is suggested to have a tumor suppressing role by degrading GI tract-specific oncogenes. CAPN11 is expressed in testis, whereas CAPN12 is expressed in hair follicles.
Among the non-classical calpains, human CAPN5 is homologous to the C. elegans sex determining gene TRA-3 and expressed in most tissues, but especially in the central nervous system. CAPN6 is highly homologous to CAPN5, is located on the X chromosome and is also suggested to be involved in sex determination. CAPN7 has a more divergent calpain sequence making it easier to design a calpain specific antigene to this member, and the divergence includes the MIT-MIT, and this calpain sequence is more similar to the fungal calpain Aspergillus nidulans. It regulates timing and completion of abscission by both the tandem MIT distinct motifs to complete cytokinesis, secure checkpoint maintenance and separate dividing cells. The human CAPN10 gene is most highly expressed in the heart, followed by the pancreas, brain, liver and kidney.
A subset of the non-canonical calpains are associated with various types of diseases and cancers. Calpain-5 can cause the rare ocular autoimmune disorder Autosomal Dominant Neovascular Inflammatory Vitreoretinopathy (ADNIV). CAPN6 has increased expression in uterine cancers, whereas CAPN9 has reduced expression in gastric cancer and is suggested to cause gastric cancer and suppression of tumorigenesis. Calpain-10 is linked to colorectal cancer, pancreatic cancer and type 2 diabetes. Mutated CAPN10 gene versions elicit a reduced rate of insulin-mediated glucose turnover, explaining how loss of calpain-10 function leads to type 2 diabetes. Increased numbers of free fatty acids in obese people leads to increased Ca2+ activation of calpain, leading to induces major endoplasmic reticulum stress markers causing cellular apoptosis. Calpain-10 is also needed for renal cell viability and decreases with age. It is further important for tubule repair in renal cells, a cleavage of cytoskeleton proteins that may lead to increased membrane permeability causing renal cell death. Multiple calpains are linked to neuronal death, apoptosis, deficits of synaptic transmission, lens cytoskeletal protein cleavage and cleavage of oncogene products.
Calpains are reported to be markers of melanoma progression such as colorectal adenocarcinomas, breast and prostate cancer, of tuberculosis, and of sudden cardiac death, and are highly expressed in melanoma cell lines and bovine bladder tumors. Like the land plant calpain DEK1, some animal calpains are connected with Ca2+ channel activity, the cytoskeleton, and/or microtubule organization. Calpain-1 and -2 bind to TRPC6 in mice and regulate the cytoskeleton and the human calpain-1 and -2 proteins cleave and activate TRPC5 associated with neuronal growth.
CAPN1, CAPN2 and CAPN6 are all reported to be involved in cytoskeletal organization and CAPN5, 6, 7 and 10 are found to stabilize MT. This is all consistent with our suggested role for calpains in regulating MTOC activity in animals as in land plants. We have linked the DEK1 calpain to controlling plants MTs through the MTOC, and in animals this is further associated with immunology and T-cell receptors (TCR) reorganizing of the MTOC linked to dynein, GTPases, integrins and actin.
Multiple human calpains are involved in cytoskeletal remodeling. Cytoskeletal connections between calpains and cancer include calpain-mediated cleavage product MYC-nick (MYC proto-oncoprotein) that promotes cytoskeletal remodeling, which is upregulated in cancer cells. Myc oncoproteins are widely involved in oncogenesis, and calpain-3 cleavage generates myc-nick that induces alpha-tubulin altered cell morphology by recruiting histone GCN5 to MT, and that drives cytoplasmic reorganization and differentiation. Accelerated calpain cleavage of the human epidermal growth factor receptor (HER2) has been found to repress the targeting of such cancers, since they contribute to resistance to anticancer therapies. Further reduction of antitumor therapy effectiveness is caused by calpain-1 and -2 modifications, since they regulate the cellular efflux machineries for drug efflux.
Calpains have similarly been suggested to cause cancer by reduced cell adhesion leading to cell release from the cellular matrix, including cell transformation, migration and invasion. Since calpains are recognized as key regulators of cell adhesion, they could promote either epithelial cell clearance or migration through adherens junctions (AJ) and focal adhesion (FA) complexes. Interestingly AJs are found crucial to epithelia identity and are further linked to the actin cytoskeleton, the establishment of polarity and cell-cell communication needed for cell proliferation and movement. RhoA and Piezo are downstream targets of FA, and are regulated by the DEK1 calpain. Both calpain-1and calpain-2 are known to target actin binding proteins, and can proteolyze E-cadherin and other adhesion proteins during mammary gland development. These data show the important role of calpains in cell adhesion disruption and actin dynamics, further suggesting roles not yet discovered.
Interestingly the function of calpains in tumor cell migration and invasion is better understood than their role in apoptosis and cancer survival, related to cell adhesion and actin dynamics. Stress is further linked to increased Ca2+ levels and overactive calpains causing endothelial cell dysfunction, increased cytoskeleton degradation and organ dysfunction implicated in cardiac dysfunction. Downregulation of several calpains are therefore suggested as therapeutic interventions in cancer treatments.
Emerging calpain-targeted therapeutic strategies are being developed to reduce calpain activities through alternatives to calpastatin regulation, including peptidyl epoxide, aldehyde and ketoamide. However, the limiting factor is the lack of calpain specificity, pointing to selecting locations outside of the shared domain sequences. Many more therapeutic options are expected as we understand calpain functions better to predict the right balance to maintain healthy cells and prevent diseases linked to calpain dysfunction.
Conclusion
Members of the calpain family of cysteine proteases are associated with a variety of severe human diseases, including cancer, multiple sclerosis and various neurological disorders. The canonical calpain cysteine protease domain arose in bacteria, followed by a massive expansion of domain structures such that extant calpain proteins consist of multiple domains in combinations that vary across the evolutionary spectrum. Correspondingly, calpains have been linked to a wide range of biological functions in plants and animals; many of these are associated with cell membranes and cytoskeletal remodeling. Such functions point to a potential core role for calpains in organizing microtubules within cells, as shown for the land plant calpain DEK1. With therapeutic strategies emerging to alter calpain activity and thus reduce cellular dysfunction, further investigation of calpain cellular and biochemical functions in animals, plants and algae can provide new directions for therapeutic intervention.
Conflicts of Interest Statement
The authors declare no conflicts of interest.
Funding Statement
J.C.F. was funded by the United States Department of Agriculture (CRIS 2030-21210-001-00D).
References
1. Murachi T, Tanaka K, Hatanaka M, Murakami T. Intracellular Ca2+-dependent protease (calpain) and its high-molecular-weight endogenous inhibitor (calpastatin). Advances in enzyme regulation. 1981;19:407-424.
2. Nian H, Ma B. Calpain–calpastatin system and cancer progression. Biological Reviews. 2021;96(3):961-975.
3. Spinozzi S, Albini S, Best H, Richard I. Calpains for dummies: What you need to know about the calpain family. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics. 2021;1869(5):140616.
4. Sorimachi H, Hata S, Ono Y. Calpain chronicle—an enzyme family under multidisciplinary characterization. Proceedings of the Japan Academy, Series B. 2011;87(6):287-327.
5. Hosfield CM, Elce JS, Davies PL, Jia Z. Crystal structure of calpain reveals the structural basis for Ca2+‐dependent protease activity and a novel mode of enzyme activation. The EMBO journal. 1999;
6. Strobl S, Fernandez-Catalan C, Braun M, et al. The crystal structure of calcium-free human m-calpain suggests an electrostatic switch mechanism for activation by calcium. Proceedings of the National Academy of Sciences. 2000;97(2):588-592.
7. Rawlings ND. Bacterial calpains and the evolution of the calpain (C2) family of peptidases. Biology Direct. 2015;10:1-12.
8. Vešelényiová D, Hutárová L, Lukáčová A, Schneiderová M, Vesteg M, Krajčovič J. Calpains in cyanobacteria and the origin of calpains. Scientific Reports. 2022;12(1):13872.
9. Goll DE, Thompson VF, Li H, Wei W, Cong J. The calpain system. Physiol Rev. Jul 2003;83(3):731-801. doi:10.1152/physrev.00029.2002
10. Zhao S, Liang Z, Demko V, et al. Massive expansion of the calpain gene family in unicellular eukaryotes. BMC Evol Biol. Sep 29 2012;12:193. doi:10.1186/1471-2148-12-193
11. Lid SE, Gruis D, Jung R, et al. The defective kernel 1 (dek1) gene required for aleurone cell development in the endosperm of maize grains encodes a membrane protein of the calpain gene superfamily. Proceedings of the National Academy of Sciences. 2002;99(8):5460-5465.
12. Denoeud F, Godfroy O, Cruaud C, et al. Evolutionary genomics of the emergence of brown algae as key components of coastal ecosystems. bioRxiv. 2024:2024.02. 19.579948.
13. Liang Z, Brown RC, Fletcher JC, Opsahl-Sorteberg H-G. Calpain-Mediated Positional Information Directs Cell Wall Orientation to Sustain Plant Stem Cell Activity, Growth and Development. Plant and Cell Physiology. 2015;56(9):1855-1866. doi:10.1093/pcp/pcv110
14. Liang Z, Demko V, Wilson RC, et al. The catalytic domain CysPc of the DEK 1 calpain is functionally conserved in land plants. The Plant Journal. 2013;75(5):742-754.
15. Araujo H, Julio A, Cardoso M. Translating genetic, biochemical and structural information to the calpain view of development. Mechanisms of Development. 2018;154:240-250.
16. Šafranek M, Shumbusho A, Johansen W, et al. Membrane-anchored calpains–hidden regulators of growth and development beyond plants? Frontiers in Plant Science. 2023;14:1289785.
17. Jékely G, Friedrich P. Characterization of two recombinant Drosophila calpains: CALPA and a novel homolog, CALPB. Journal of Biological Chemistry. 1999;274(34):23893-23900.
18. Wang C, Barry JK, Min Z, Tordsen G, Rao AG, Olsen O-A. The calpain domain of the maize DEK1 protein contains the conserved catalytic triad and functions as a cysteine proteinase. Journal of Biological Chemistry. 2003;278(36):34467-34474.
19. Gibbons IR, Rowe AJ. Dynein: a protein with adenosine triphosphatase activity from cilia. Science. 1965;149(3682):424-426.
20. Tran D, Galletti R, Neumann ED, et al. A mechanosensitive Ca2+ channel activity is dependent on the developmental regulator DEK1. Nature Communications. 2017;8(1):1009.
21. Kaczmarek JS, Riccio A, Clapham DE. Calpain cleaves and activates the TRPC5 channel to participate in semaphorin 3A-induced neuronal growth cone collapse. Proceedings of the National Academy of Sciences. 2012;109(20):7888-7892.
22. Arias-Darraz L, Cabezas D, Colenso CK, et al. A transient receptor potential ion channel in Chlamydomonas shares key features with sensory transduction-associated TRP channels in mammals. The Plant Cell. 2015;27(1):177-188.
23. Owsianik G, D’hoedt D, Voets T, Nilius B. Structure–function relationship of the TRP channel superfamily. Reviews of physiology, biochemistry and pharmacology. 2006:61-90.
24. Hellwig N, Albrecht N, Harteneck C, Schultz Gn, Schaefer M. Homo-and heteromeric assembly of TRPV channel subunits. Journal of cell science. 2005;118(5):917-928.
25. Himmel NJ, Cox DN. Transient receptor potential channels: current perspectives on evolution, structure, function and nomenclature. Proceedings of the Royal Society B. 2020;287(1933):20201309.
26. Lid SE, Olsen L, Nestestog R, et al. Mutation in the Arabidopisis thaliana DEK1 calpain gene perturbs endosperm and embryo development while over-expression affects organ development globally. Planta. 2005;221:339-351.
27. Tian Q, Olsen L, Sun B, et al. Subcellular localization and functional domain studies of DEFECTIVE KERNEL 1 in maize and Arabidopsis suggest a model for aleurone cell fate specification involving CRINKLY4 and SUPERNUMERARY ALEURONE LAYER1. The Plant Cell. 2007;19(10):3127-3145. doi:10.1105/tpc.106.048868
28. Perroud PF, Demko V, Johansen W, Wilson RC, Olsen OA, Quatrano RS. Defective Kernel 1 (DEK 1) is required for three‐dimensional growth in P hyscomitrella patens. New Phytologist. 2014;203(3):794-804.
29. Liu Y, Li X, Zhao J, et al. Direct evidence that suspensor cells have embryogenic potential that is suppressed by the embryo proper during normal embryogenesis. Proc Natl Acad Sci U S A. Oct 6 2015;112(40):12432-7. doi:10.1073/pnas.1508651112
30. Friml J, Vieten A, Sauer M, et al. Efflux-dependent auxin gradients establish the apical–basal axis of Arabidopsis. Nature. 2003;426(6963):147-153.
31. Perroud PF, Meyberg R, Demko V, Quatrano RS, Olsen OA, Rensing SA. DEK1 displays a strong subcellular polarity during Physcomitrella patens 3D growth. New phytologist. 2020;226(4):1029-1041.
32. Yan Y, Sun Z, Yan P, Wang T, Zhang Y. Mechanical regulation of cortical microtubules in plant cells. New Phytologist. 2023;239(5):1609-1621.
33. Jánossy J, Ubezio P, Apáti Á, Magócsi M, Tompa P, Friedrich P. Calpain as a multi-site regulator of cell cycle. Biochemical pharmacology. 2004;67(8):1513-1521.
34. Franco SJ, Huttenlocher A. Regulating cell migration: calpains make the cut. Journal of cell science. 2005;118(17):3829-3838.
35. Santos DM, Xavier JM, Morgado AL, Sola S, Rodrigues CM. Distinct regulatory functions of calpain 1 and 2 during neural stem cell self-renewal and differentiation. PloS one. 2012;7(3):e33468.
36. Potz BA, Abid MR, Sellke FW. Role of calpain in pathogenesis of human disease processes. Journal of nature and science. 2016;2(9)
37. Shapovalov I, Harper D, Greer PA. Calpain as a therapeutic target in cancer. Expert Opinion on Therapeutic Targets. 2022;26(3):217-231.
38. Ferreira A. Calpain dysregulation in Alzheimer’s disease. International Scholarly Research Notices. 2012;2012
39. Mahaman YAR, Huang F, Wu M, et al. Moringa oleifera alleviates homocysteine-induced Alzheimer’s disease-like pathology and cognitive impairments. Journal of Alzheimer’s Disease. 2018;63(3):1141-1159.
40. Shams R, Banik NL, Haque A. Calpain in the cleavage of alpha-synuclein and the pathogenesis of Parkinson’s disease. Progress in molecular biology and translational science. 2019;167:107-124.
41. Gafni J, Ellerby LM. Calpain activation in Huntington’s disease. Journal of Neuroscience. 2002;22(12):4842-4849.
42. Rao SS, Mu Q, Zeng Y, et al. Calpain‐activated mTORC2/Akt pathway mediates airway smooth muscle remodelling in asthma. Clinical & Experimental Allergy. 2017;47(2):176-189.
43. Tabata C, Tabata R, Nakano T. The calpain inhibitor calpeptin prevents bleomycin-induced pulmonary fibrosis in mice. Clinical & Experimental Immunology. 2010;162(3):560-567.
44. Ono Y, Saido TC, Sorimachi H. Calpain research for drug discovery: challenges and potential. Nature Reviews Drug Discovery. 2016;15(12):854-876.
45. Carafori E. Calpain: a protease in search of a function? Biochem Biophys Res Commun. 1998;247:193-203.
46. Azam M, Andrabi SS, Sahr KE, Kamath L, Kuliopulos A, Chishti AH. Disruption of the mouse μ-calpain gene reveals an essential role in platelet function. Molecular and cellular biology. 2001;21(6):2213-2220.
47. Gan-Or Z, Bouslam N, Birouk N, et al. Mutations in CAPN1 cause autosomal-recessive hereditary spastic paraplegia. The American Journal of Human Genetics. 2016;98(5):1038-1046.
48. Smith AW, Das A, Guyton MK, Ray SK, Rohrer B, Banik NL. Calpain inhibition attenuates apoptosis of retinal ganglion cells in acute optic neuritis. Investigative Ophthalmology & Visual Science. 2011;52(7):4935-4941.
49. Wang KK. Calpain and caspase: can you tell the difference? Trends in neurosciences. 2000;23(1):20-26.
50. Wang Y, Liu Y, Bi X, Baudry M. Calpain-1 and calpain-2 in the brain: new evidence for a critical role of calpain-2 in neuronal death. Cells. 2020;9(12):2698.
51. Morton JD, Lee HY, McDermott JD, et al. A macrocyclic calpain inhibitor slows the development of inherited cortical cataracts in a sheep model. Investigative ophthalmology & visual science. 2013;54(1):389-395.
52. Huang Y, Wang KK. The calpain family and human disease. Trends in molecular medicine. 2001;7(8):355-362.
53. Conze C, Rierola M, Trushina NI, et al. Caspase-cleaved tau is senescence-associated and induces a toxic gain of function by putting a brake on axonal transport. Molecular psychiatry. 2022;27(7):3010-3023.
54. Azuma M, Fukiage C, David L, Shearer T. Activation of calpain in lens: a review and proposed mechanism. Experimental eye research. 1997;64(4):529-538.
55. Richard I, Broux O, Allamand V, et al. Mutations in the proteolytic enzyme calpain 3 cause limb-girdle muscular dystrophy type 2A. Cell. 1995;81(1):27-40.
56. Sorimachi H, Imajoh-Ohmi S, Emori Y, et al. Molecular cloning of a novel mammalian calcium-dependent protease distinct from both m-and μ-types: specific expression of the mRNA in skeletal muscle. Journal of Biological Chemistry. 1989;264(33):20106-20111.
57. Moretti D, Del Bello B, Allavena G, Corti A, Signorini C, Maellaro E. Calpain-3 impairs cell proliferation and stimulates oxidative stress-mediated cell death in melanoma cells. PloS one. 2015;10(2):e0117258.
58. Roperto S, De Tullio R, Raso C, et al. Calpain3 is expressed in a proteolitically active form in papillomavirus-associated urothelial tumors of the urinary bladder in cattle. PLoS One. 2010;5(4):e10299.
59. Kramerova I, Kudryashova E, Wu B, Ottenheijm C, Granzier H, Spencer MJ. Novel role of calpain-3 in the triad-associated protein complex regulating calcium release in skeletal muscle. Human molecular genetics. 2008;17(21):3271-3280.
60. Peng P, Wu W, Zhao J, et al. Decreased expression of Calpain-9 predicts unfavorable prognosis in patients with gastric cancer. Scientific reports. 2016;6(1):29604.
61. Matena K, Boehm T, Dear TN. Genomic organization of mouse Capn5 and Capn6 Genes confirms that they are a distinct calpain subfamily. Genomics. 1998;48(1):117-120.
62. Franz T, Vingron M, Boehm T, Dear TN. Capn7: a highly divergent vertebrate calpain with a novel C-terminal domain. Mammalian genome. 1999;10:318-321.
63. Paine EL, Skalicky JJ, Whitby FG, et al. The Calpain-7 protease functions together with the ESCRT-III protein IST1 within the midbody to regulate the timing and completion of abscission. bioRxiv. 2022:2022.10.18.512775. doi:10.1101/2022.10.18.512775
64. Storr SJ, Carragher NO, Frame MC, Parr T, Martin SG. The calpain system and cancer. Nature Reviews Cancer. 2011;11(5):364-374.
65. Tang PH, Chemudupati T, Wert KJ, et al. Phenotypic variance in Calpain-5 retinal degeneration. American journal of ophthalmology case reports. 2020;18:100627.
66. Frances CP, Conde MC, Saez ME, et al. Identification of a protective haplogenotype within CAPN10 gene influencing colorectal cancer susceptibility. Journal of gastroenterology and hepatology. 2007;22(12):2298-2302.
67. Fong P-y, Fesinmeyer MD, White E, et al. Association of diabetes susceptibility gene calpain-10 with pancreatic cancer among smokers. Journal of gastrointestinal cancer. 2010;41:203-208.
68. Horikawa Y, Oda N, Cox NJ, et al. Genetic variation in the gene encoding calpain-10 is associated with type 2 diabetes mellitus. Nature genetics. 2000;26(2):163-175.
69. Liu X, Schnellmann RG. Calpain mediates progressive plasma membrane permeability and proteolysis of cytoskeleton-associated paxillin, talin, and vinculin during renal cell death. Journal of Pharmacology and Experimental Therapeutics. 2003;304(1):63-70.
70. Frangié C, Zhang W, Perez J, Dubois Y-CX, Haymann J-P, Baud L. Extracellular calpains increase tubular epithelial cell mobility: implications for kidney repair after ischemia. Journal of biological chemistry. 2006;281(36):26624-26632.
71. Liu K, Li L, Cohen SN. Antisense RNA-mediated deficiency of the calpain protease, nCL-4, in NIH3T3 cells is associated with neoplastic transformation and tumorigenesis. Journal of Biological Chemistry. 2000;275(40):31093-31098.
72. Shiba E, Kambayashi J-I, Sakon M, et al. Ca 2+-dependent neutral protease (Calpain) activity in breast cancer tissue and estrogen receptor status. Breast Cancer. 1996;3:13-17.
73. Kulkarni S, Reddy K, Esteva F, Moore H, Budd G, Tubbs R. Calpain regulates sensitivity to trastuzumab and survival in HER2-positive breast cancer. Oncogene. 2010;29(9):1339-1350.
74. Hong JY, Park SY, Kim Y, Lee CY, Lee MG. Calpain and spectrin breakdown products as potential biomarkers in tuberculous pleural effusion. Journal of Thoracic Disease. 2018;10(5):2558.
75. Kunišek L, Ilijaš KM, Medved I, et al. Cardiomyocytes calpain 2 expression: Diagnostic forensic marker for sudden cardiac death caused by early myocardial ischemia and an indicator of the duration of myocardial agonal period? Medical hypotheses. 2022;158:110738.
76. Farmer LK, Rollason R, Whitcomb DJ, et al. TRPC6 binds to and activates calpain, independent of its channel activity, and regulates podocyte cytoskeleton, cell adhesion, and motility. Journal of the American Society of Nephrology. 2019;30(10):1910-1924.
77. Lasserre R, Alcover A. Cytoskeletal cross-talk in the control of T cell antigen receptor signaling. FEBS letters. 2010;584(24):4845-4850.
78. Conacci-Sorrell M, Ngouenet C, Eisenman RN. Myc-nick: a cytoplasmic cleavage product of Myc that promotes α-tubulin acetylation and cell differentiation. Cell. 2010;142(3):480-493.
79. Leloup L, Wells A. Calpains as potential anti-cancer targets. Expert opinion on therapeutic targets. 2011;15(3):309-323.
80. García-Trevijano ER, Ortiz-Zapater E, Gimeno A, Viña JR, Zaragozá R. Calpains, the proteases of two faces controlling the epithelial homeostasis in mammary gland. Frontiers in Cell and Developmental Biology. 2023;11:1249317.
81. Pardo-Pastor C, Rubio-Moscardo F, Vogel-González M, et al. Piezo2 channel regulates RhoA and actin cytoskeleton to promote cell mechanobiological responses. Proceedings of the National Academy of Sciences. 2018;115(8):1925-1930.
82. Fernández LR. Subcellular Distribution of Calpain-1 and Calpain-2 as a Key Event for Calpain-mediated Functions in Physiological and Neoplastic Mammary Models. Universitat de València. Facultat de Ciències Biològiques.; 2019.
83. Demarchi F, Bertoli C, Greer P, Schneider C. Ceramide triggers an NF-κB-dependent survival pathway through calpain. Cell Death & Differentiation. 2005;12(5):512-522.
84. Li C, Chen S, Yue P, et al. Proteasome inhibitor PS-341 (bortezomib) induces calpain-dependent IκBα degradation. Journal of Biological Chemistry. 2010;285(21):16096-16104.
85. Carragher NO. Calpain inhibition: a therapeutic strategy targeting multiple disease states. Current pharmaceutical design. 2006;12(5):615-638.
86. Dong B, Liu R. Characterization of endogenous and recombinant human calpain-10. Biochimie. 2008;90(9):1362-1371.
87. Pánico P, Salazar AM, Burns AL, Ostrosky-Wegman P. Role of calpain-10 in the development of diabetes mellitus and its complications. Archives of medical research. 2014;45(2):103-115.
88. Ben‐Aharon I, Brown PR, Shalgi R, Eddy EM. Calpain 11 is unique to mouse spermatogenic cells. Molecular Reproduction and Development: Incorporating Gamete Research. 2006;73(6):767-773.
89. Charlton R, Henderson M, Richards J, et al. Immunohistochemical analysis of calpain 3: advantages and limitations in diagnosing LGMD2A. Neuromuscular Disorders. 2009;19(7):449-457.
90. Hauerslev S, Sveen M-L, Duno M, Angelini C, Vissing J, Krag TO. Calpain 3 is important for muscle regeneration: evidence from patients with limb girdle muscular dystrophies. BMC Musculoskeletal Disorders. 2012;13:1-11.
91. Mahajan VB, Skeie JM, Bassuk AG, et al. Calpain-5 mutations cause autoimmune uveitis, retinal neovascularization, and photoreceptor degeneration. PLoS Genetics 2012; 8(10): e1003001.
92. Turner MD, Cassell PG, Hitman GA. Calpain‐10: from genome search to function. Diabetes/metabolism research and reviews. 2005;21(6):505-514.
93. Zha C, Farah CA, Holt RJ, et al. Biallelic variants in the small optic lobe calpain CAPN15 are associated with congenital eye anomalies, deafness and other neurodevelopmental deficits. Human molecular genetics. 2020;29(18):3054-3063.
94. Litosh VA, Rochman M, Rymer JK, Porollo A, Kottyan LC, Rothenberg ME. Calpain-14 and its association with eosinophilic esophagitis. Journal of Allergy and Clinical Immunology. 2017;139(6):1762-1771. e7.