





Crosstalk Between Adipocytes and Endothelial Cells in Angiolipoma
Adipocyte-Endothelial Cell Crosstalk in Angiolipoma: Contribution of the Extracellular Galectin-1, -3 and -8 and their Binding Partners
Enrique Arciniegas¹*, Andrés Duque¹, Héctor Rojas², Jacinto Pineda³, Antonio Salgado⁴, Alejandra Urbano⁵, Ana María Sáenz⁵, Richard Ramírez⁴
- Institute of Biomedicine, Central University of Venezuela, Caracas, Venezuela.
- Institute of Immunology, Central University of Venezuela, Caracas, Venezuela.
- Institute of Anatomy and Pathology, Central University of Venezuela, Caracas, Venezuela.
- Autonomous Service Institute of Biomedicine, Caracas, Venezuela.
- Service of Dermatology, University Hospital of Caracas, Venezuela.
OPEN ACCESS
PUBLISHED: 31 March 2026
CITATION: Arciniegas, E., Duque, A., et al., 2026. Adipocyte-Endothelial Cell Crosstalk in Angiolipoma: Contribution of the Extracellular Galectin-1, -3 and -8 and their Binding Partners. Medical Research Archives, [online] 14(3).
COPYRIGHT: © 2026 European Society of Medicine. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
ISSN 2375-1924
Abstract
Angiolipomas are benign subcutaneous nodules composed of mature adipose tissue with thin-walled, dilated and tortuous small blood vessels, displaying intraluminal deposition of fibrin microthrombi formed from fibrinogen. The latter, considered not only as a diagnostic hallmark of angiolipoma, but also as a marker of vascular permeability. It is known that human adipose tissue (AT) growth, expansion, and maintenance is angiogenesis dependent and involves adipocytes, immune cells, fibroblasts, adipose stem cells and preadipocytes, all of them supported by an extracellular matrix. Interestingly, emerging evidence have emphasized that adipocytes maintain a bidirectional communication (crosstalk) with adipose endothelial cells (ECs) in healthy and pathological tissues and that this reciprocal and dynamic communication is crucial not only for the growth, expansion and maintenance of the AT, but also for the formation of new capillaries (angiogenesis) or microvasculature remodeling. However, the signals and mediators involved in this communication remain to be investigated. Herein, we show that in angiolipoma GAL-1, -3 and -8 and some of their binding partners that includes proteoglycans such as perlecan, agrin, CD44 and endocan, as well as glycoproteins including TLR4, fibronectin, thrombospondin-1, integrin β1, PECAM-1, endoglin, VE-cadherin, and neuropilin-1, were immunolocalized around mature adipocytes that were in close proximity to small blood vessels, as well as in the ECs lining the lumen of these vessels. We propose that in angiolipoma, GAL-1, -3 and -8 acting in concert with their binding partners form a supramolecular multivalent structure on the surface of mature adipocytes and ECs that might be facilitating the bidirectional communication (crosstalk) between adipocytes and ECs, contributing to the mature adipocytic expansion and the new vasculature formation (angiogenesis) or microvasculature remodeling.
Keywords
- Angiolipoma
- adipocyte
- endothelial cell
- crosstalk
- galectins
- Gal-glycan lattice
Introduction
Angiolipomas are benign subcutaneous nodules composed of mature adipose tissue with thin-walled, dilated and tortuous small blood vessels, displaying intraluminal deposition of fibrin microthrombi formed from fibrinogen. The latter, considered not only as a diagnostic hallmark of angiolipoma, but also as a marker of vascular permeability. Equally well established is that human adipose tissue (AT) is comprised mainly of mature adipocytes with every adipocyte surrounded by one or more capillaries and that AT growth, expansion, and maintenance is angiogenesis dependent and involves adipocytes, immune cells, fibroblasts, adipose stem cells and preadipocytes. All of them supported by an extracellular matrix whose remodeling has been suggested is required for healthy AT expansion. Respect to AT growth, expansion and angiogenesis, in vitro studies revealed that AT explants in fibrin, collagen or matrigel, trigger blood vessel formation and that adipose ECs in turn promote preadipocyte differentiation, suggesting that the adipose ECs may be involved in the positive regulation of growth and maintenance of AT through their direct adhesion to mature adipocytes. Interestingly, emerging evidence have emphasized that adipocytes maintain a bidirectional communication (crosstalk) with adipose ECs in healthy and pathological tissues via endocrine and paracrine signals and that this reciprocal and dynamic communication is crucial not only for the growth, expansion and maintenance of the AT, but also for the formation of new capillaries (angiogenesis) or microvasculature remodeling. In this context, models of co-culture adipocyte-EC have been developed to study the crosstalk of mature adipocytes and ECs. However, the signals and mediators involved in this communication remain to be investigated. Of relevance, interplay between adipocyte and adipose EC during the formation of capillary structures involving the production of angiogenic factors by adipocytes, particularly vascular endothelial growth factor (VEGF), and the activation of their receptors tyrosine kinases (VEGFR1/VEGFR2) present on ECs, is well documented. Moreover, dynamic communication between adipocytes and ECs involving heparan sulfate proteoglycans (HSPGs) (e.g., perlecan, agrin, syndecan-1 (CD138) and chondroitin sulfate proteoglycans (CSPGs) (e.g., CD44, betaglycan) that surround both adipocytes and vascular structures has been recently reported. In addition, it has been suggested that paracrine activities exerted by free soluble proteoglycans as endocan, a small soluble dermatan sulfate proteoglycan (DSPG) (also known as endothelial cell specific molecule-1, ESM-1) expressed by adipocytes and activated capillary endothelial cells, are highly regulated by pro-angiogenic molecules such as VEGF and fibroblast growth factor (FGF). Likewise, accumulating evidence have suggested that ECs and mature adipocytes express Toll-like receptor (TLR) 2 and TLR4 on their surface and that both TLRs are activated by paracrine endogenous ligands that include fibrinogen, HSPGs, fibronectin (FN) and galectin-3, contributing to EC activation, formation of microvasculature, and inflammation as well as adipocyte differentiation. Thus, it is possible that both TLRs could be also participating in the crosstalk between adipocytes and ECs. Another molecules that could be controlling this bidirectional communication in AT, are the soluble proteins family named galectins (GALs), particularly Galectin-1 (GAL-1), GAL-3, and GAL-8. GALs are a highly conserved family of 16 different small proteins identified in mammals. They are characterized by their ability to bind exposed β-galactosides motifs, particularly lactosamine (LacNAC) sequences in glycoconjugates and defined by the presence of one or two conserved Carbohydrate Recognition Domains (CRDs). Individual GAL family members have been involved in many biological processes including cell-cell communication (crosstalk), cell-ECM interaction and angiogenesis. Importantly, GALs function extracellularly and intracellularly. Extracellular GALs can appear arranged into non-covalent bound dimers and oligomeric forms able to bind and cross-link specific glycoconjugates containing N- and/or O-glycan residues or appear organized in supramolecular multivalent complexes known as galectin-glycan (GAL-glycan) lattices on the cell surface and in the extracellular milieu. The GAL-glycan lattices can mediate, through specific signaling pathways, cellular processes including cell-cell communication (crosstalk), cell matrix adhesion, proliferation, growth, differentiation, migration, and inflammation. Nevertheless, the current knowledge about how the aforementioned protein interactors would contribute to the bidirectional communication between adipocytes and AT endothelium, particularly in angiolipoma, is still limited. Thus, we believe that to understanding the interactions and contribution of these proteins to the crosstalk between adipocyte and EC in healthy and pathological tissues, including in angiolipoma, will be necessary to define which molecules could be participating. In view of the above considerations, the purpose of this study was examine not only the presence and localization of GAL-1, -3, and -8 in angiolipoma tissue, but also consider the possibility that these GALs, acting in concert with some of their binding partners present on the adipocyte surface and on the abluminal and luminal sides of ECs, could be modulating the crosstalk between adipocyte and EC through the generation of lattices. Binding partners that contain N- and /or O-linked glycans in the extracellular domains and that include proteoglycans such as perlecan, agrin, endocan, and cluster of differentiation (CD) 44 (CD44), as well as glycoproteins such as TLR4, fibronectin (FN), thrombospondin-1 (TSP-1), integrin β1, Platelet Endothelial Cell Adhesion Molecule-1 (PECAM-1, also known as CD31), endoglin (CD105), vascular endothelial cadherin (VE-cadherin), VEGFR2, and neuropilin-1 (NRP1).
Methods
Tissue samples
Eight biopsies were obtained from patients clinically and histopathologically (H&E staining) diagnosed with angiolipoma. Data regarding gender, age and site of lesion appear listed in Table 1. This study was approved by the Ethics Committee of the Universitary Hospital of Caracas and performed in line with the principles of the Declaration of Helsinki.
| Patient | Gender | Age | Site of lesion |
|---|---|---|---|
| 1 | Female | 41 years | Thigh |
| 2 | Male | 62 years | Thigh |
| 3 | Female | 48 years | Arm |
| 4 | Male | 24 years | Arm |
| 5 | Male | 38 years | Wrist |
| 6 | Female | 48 years | Scapular region |
| 7 | Male | 28 years | Arm |
| 8 | Male | 56 years | Arm |
Indirect immunofluorescence staining of tissue sections
For each case, 4µm sections were cut from tissue blocks of biopsies that were previously fixed with formalin and embedded in paraffin. Sections were then set onto silanised glass slides. In order to remove the paraffin and to rehydrate the sections, they were first immersed in xylene, followed by a graded ethanol serie (from 100% to 70%), and rehydrated in distilled water and equilibrated in phosphate-buffered saline (PBS) for 10 min. Prior to incubation with the primary antibodies, sections were blocked for 1 hour in a humid chamber with phosphate-buffered saline (PBS) containing 3% BSA and 0.1% Tween 20, in order to prevent non-specific staining. Sections were then incubated with the primary antibodies diluted in PBS, 3% BSA, 0.1% Tween 20 for one hour at RT. Then, the sections were washed twice with PBS and incubated with anti-rabbit IgG Cruz Fluor 594 diluted with PBS containing 3% BSA and 0.1% Tween20 for 30 min in a humid chamber in the dark. Finally, the sections were washed in PBS and mounted with glass coverslips using mounting medium containing 4´,6-diamidino-2-phenylindole (DAPI) for 15 min in the dark. All images were captured using a 1X81 Olympus inverted microscope with the Fluo View Confocal Laser Scanning configuration (CLSM) equipped with software program FV10.ASW. For autofluorescence, selected sections of 4µm dewaxed and without staining were examined with the CLSM. Autofluorescence was determined from the ratio between the emission fluorescence in the wavelength range between 603 and 646nm and the excitation light with a wavelength of between 491 and 592nm. To validate antibody binding specificity, some sections were incubated with only primary antibody or secondary antibody.
Results
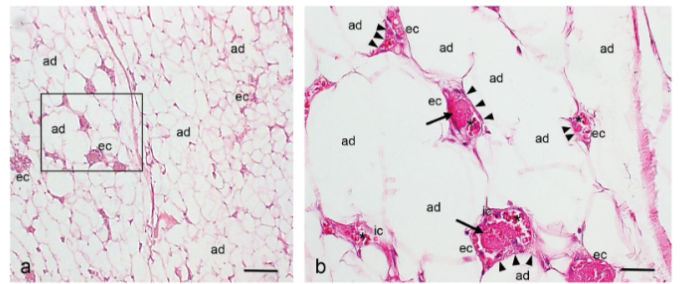
Histopathological analysis of the angiolipoma tissue sections showed the mature adipose tissue (AT) with a variable number of dilated and tortuous small blood vessels containing fibrin microthrombi, red blood cells, and immune cells.
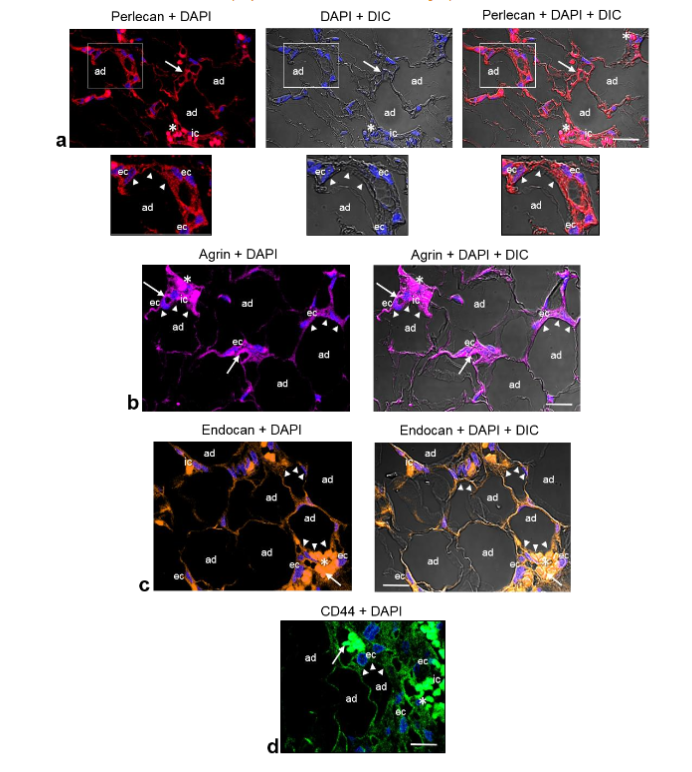
Immunolocalization of perlecan, agrin, endocan and CD44 in angiolipoma. Because the proteoglycans perlecan, agrin, endocan and CD44 are heavily glycosylated proteins that comprise a core protein and one or more covalently attached linear and heterogeneous sulfated glycans and that they surround both adipocytes and vascular structures regulating the communication between adipocytes and ECs (crosstalk), we examined the presence and localization of these proteoglycans in the angiolipoma tissue sections selected. Immunofluorescence staining analyzed by confocal microscopy revealed that perlecan, agrin, endocan, and CD44 were present around the mature adipocytes that were in close proximity to small blood vessels. In addition, moderate immunoreactivity of these proteoglycans was also observed in the endothelial cells lining the lumen of these vessels, as well as in some immune cells.
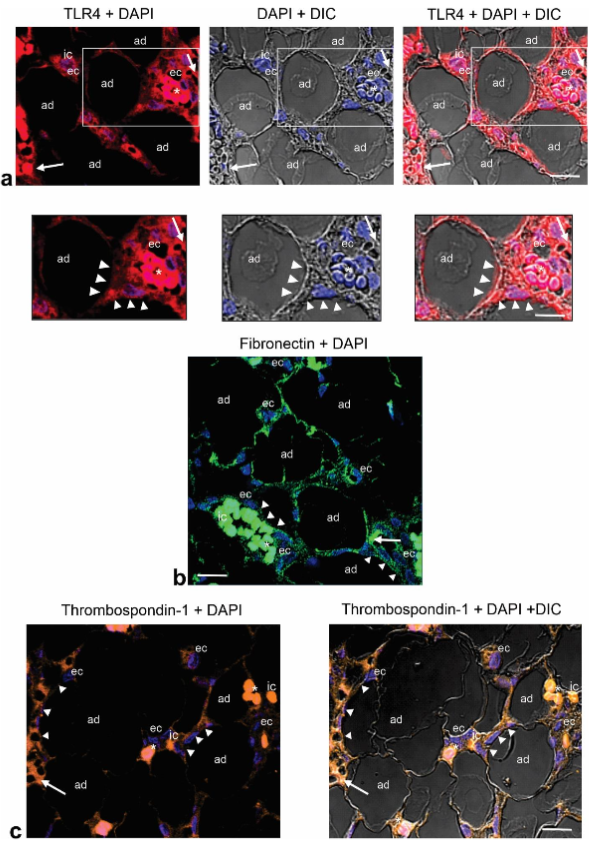
Immunolocalization of TLR4, fibronectin, and thrombospondin-1 in angiolipoma. We next examined the presence and localization of TLR4, fibronectin (FN), and thrombospondin-1 (TSP-1), glycoproteins that contain N-linked glycans and have been associated with angiogenesis and adipogenesis. Examination of angiolipoma tissue sections selected revealed an intensive TLR4 immunoreactivity around the surface of mature adipocytes that were in close proximity to small blood vessels. For FN and TSP-1, less intense immunoreactivity was observed. Both were also observed around the mature adipocytes that were proximate to small blood vessels. Additional immunostaining for TLR4, FN and TSP-1, was also evidenced in ECs lining the lumen of these vessels, as well as in some immune cells.
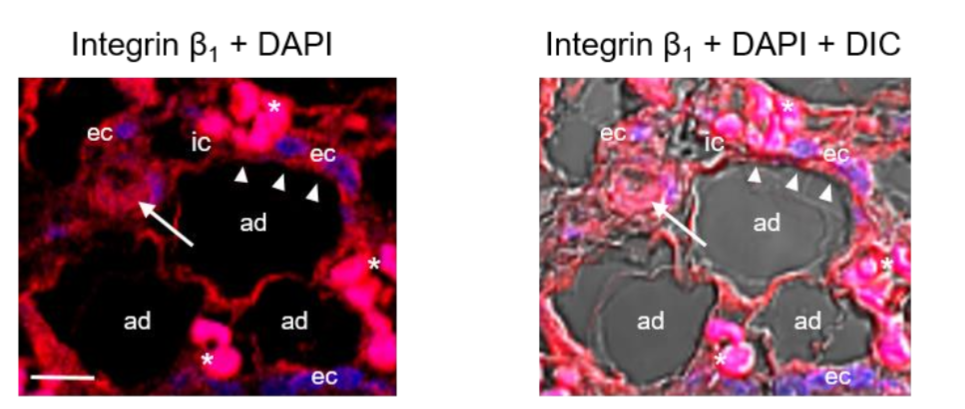
Immunolocalization of integrin β1 in angiolipoma. Integrins contain N-linked glycans and are cell-surface adhesion molecules that bind to some specific extracellular matrix molecules including FN, TSP-1, and perlecan, and have been involved in adipogenesis, and in angiogenesis. We have therefore examined the presence and localization of integrin β1 subunit in the angiolipoma tissue sections selected. Immunolocalization with anti-integrin β1 revealed that this subunit was localized around the mature adipocytes that were in close proximity to small blood vessels, as well as in the ECs lining the lumen of these vessels and some immune cells.
Immunostaining of PECAM-1/CD31, endoglin (CD105), VE-cadherin and VEGFR2 in angiolipoma. PECAM-1/CD31 is a transmembrane glycoprotein highly expressed on the surface of activated ECs that plays a critical role in the regulation of angiogenesis and vascular remodeling. It has an N-terminal ectodomain containing N-linked glycans residues, playing an important role in cell adhesion through homophilic interaction (that is, with PECAM-1) and heterophilic interactions with other glycoproteins including endoglin, VE-cadherin and VEGFR2, and proteoglycans such as CD44 and endocan. Accordingly, we examined the presence and distribution of these glycoproteins in the small blood vessels of the angiolipoma tissue sections selected. Immunofluorescence showed that PECAM-1, endoglin, VE-cadherin and VEGFR2 were localized in the ECs of the tortuous and dilated small blood vessels. Likewise, moderate PECAM-1 and endoglin immunoreactivities were observed in some immune cells.
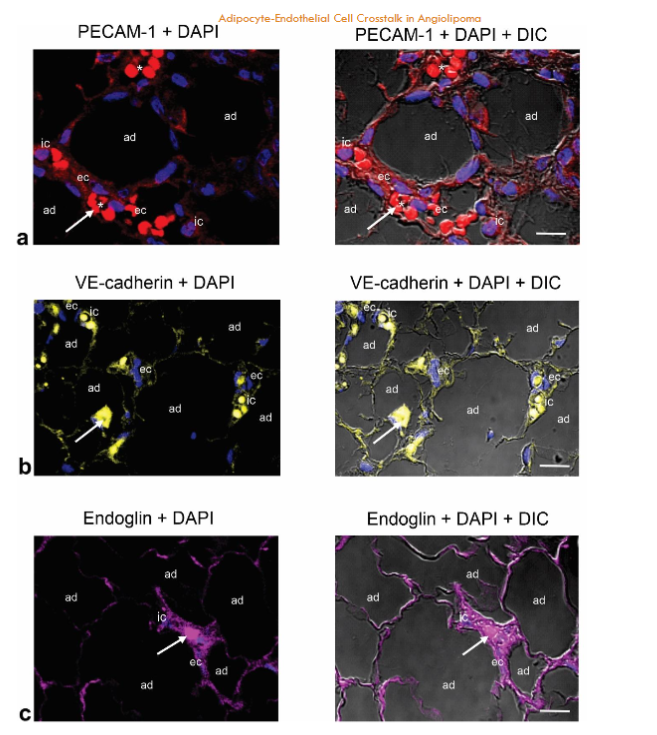
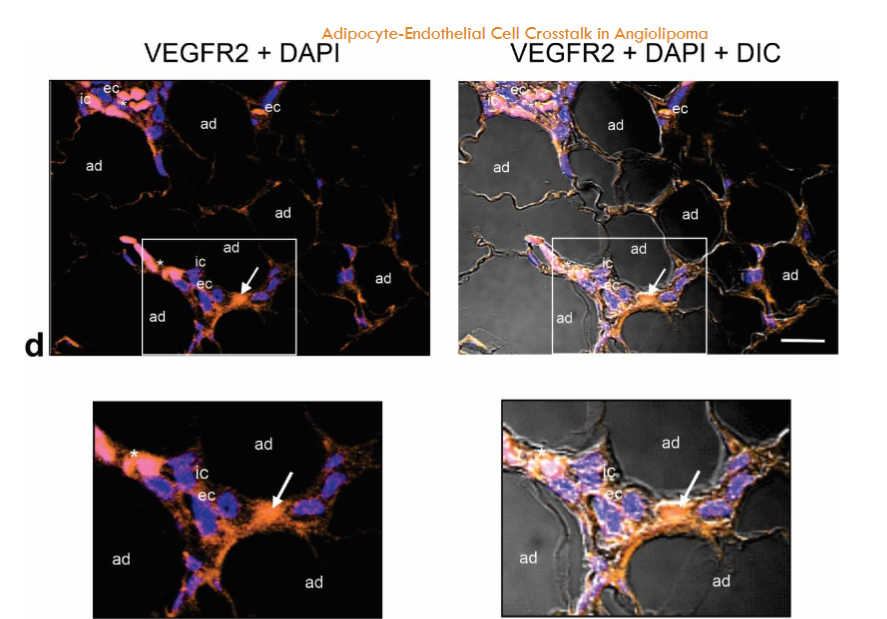
Fig. 5a-d. Immunostaining of PECAM-1 (CD31), VE-cadherin, endoglin, and VEGFR2 in the same biopsy (thigh). PECAM-1 (CD31) (a), endoglin (b), VE-cadherin (c) and VEGFR2 (d) are located in the ECs (ec) lining the lumen of the tortuous and dilated small blood vessels containing intraluminal microthrombi (arrows). Moderate PECAM-1 (CD31) and endoglin immunostaining is also observed in some immune cells (ic). Enlargement show the immunoreactivity for VEGFR2 in the ECs (ec). Blue, nuclear DAPI staining; DIC, differential interference contrast image; red blood cells (asterisk). Scale bar: 25µm (a); 35µm (b,c); 35µm, 20µm (d).
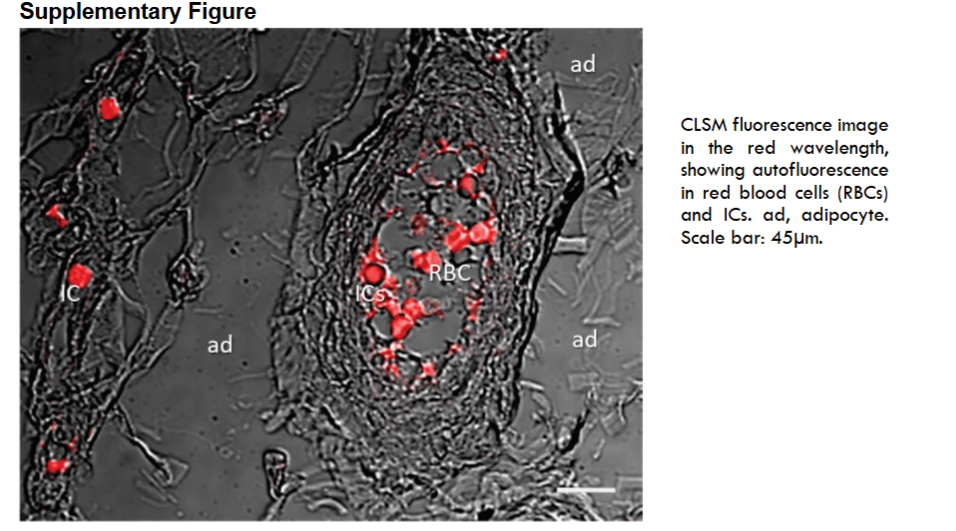
Overall, exacerbated immunopositivity in RBCs, ICs, and microthrombi provoked by autofluorescence at the red wavelength was observed in the selected sections (to see Methods section and supplementary File).
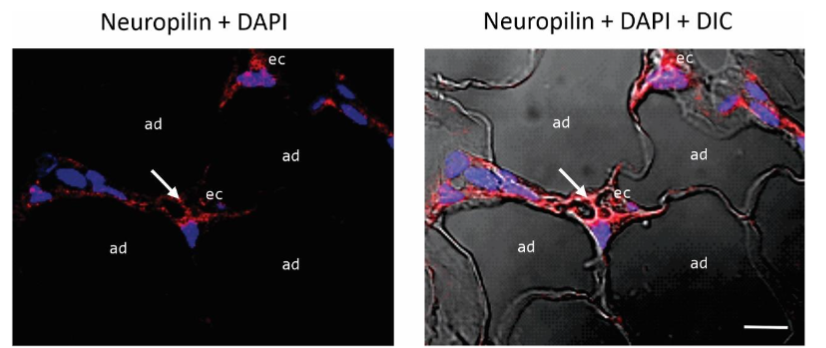
Immunolocalization of Neuropilin-1 in angiolipoma. Neuropilin-1 (NRP1) is a transmembrane glycoprotein expressed by several cell types including ECs. It is considered as a co-receptor for VEGFR2 facilitating VEGF binding and thus promoting the angiogenesis process. We have therefore examined the presence and distribution of this glycoprotein in the angiolipoma tissue sections selected. Immunofluorescence showed that NRP1 was localized in the ECs lining the lumen of tortuous and dilated small blood vessels.
Immunostaining of GAL-1, GAL-3 and GAL-8 in angiolipoma. Given that GAL-1, GAL-3, and GAL-8 bind N- and/or O-linked glycan residues and that they not only promotes EC activation, angiogenesis and vascular remodeling, but also might play an important role in AT, modulating adipocyte proliferation and differentiation, we next examined the localization and distribution of these galectins in the angiolipoma tissues selected. Immunofluorescence revealed an intensive immunoreactivity for GAL-1, -3 and -8 in the mature adipocytes that were in close proximity to tortuous and dilated small blood vessels, as well in the ECs lining the lumen of these vessels. Additional immunostaining was also evidenced in some immune cells.
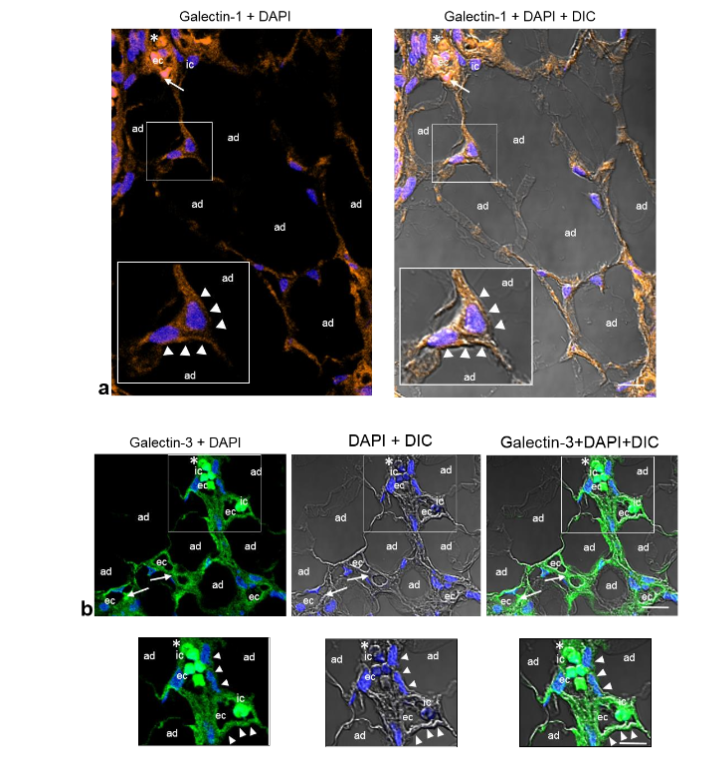
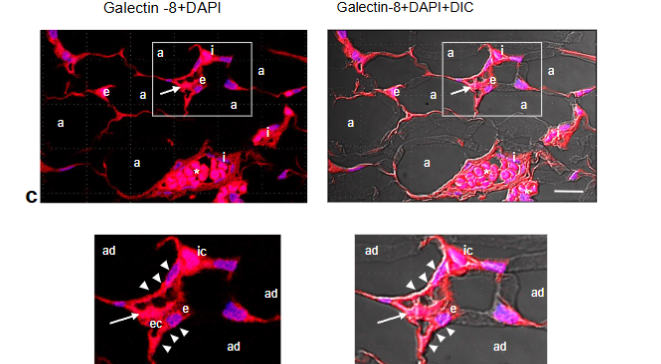
Fig. 7a-c. Representative CLSM fluorescence images of GAL-1, GAL-3, and GAL-8 in the same biopsy (thigh). GAL-1 (a), GAL-3 (b) and GAL-8 (c) are located around the mature adipocytes (ad) that are in close proximity to dilated and tortuous small blood vessels containing fibrin microthrombi (arrows), as well as in the ECs (ec) lining the vessel lumen and some immune cells (ic). Enlargements show the corresponding immunoreactivities in the mature adipocytes (ad) in close proximity to the small blood vessels (arrowheads), ECs (ec), and immune cells (ic). Blue, nuclear DAPI staining; DIC, differential interference contrast image; red blood cells (asterisk). Scale bars: 35µm, 20µm (a); 35µm, 25µm (b); 35µm, 20µm (c).
Discussion
In this study we showed that in angiolipoma tissues, the proteoglycans perlecan, agrin, CD44, and endocan were immunolocalized around mature adipocytes that were in close proximity to small blood vessels (arterioles, capillaries and venules), as well as in the ECs lining the lumen of these vessels and some immune cells. Remarkably, recent studies in AT have revealed the presence of HSPGs such as perlecan, agrin, CSPGs including CD44, and DSPG endocan, suggesting that these proteoglycans play an important role in the development and functioning of AT mediating the dynamic bidirectional communication (crosstalk) between adipocytes and microvascular ECs, promoting the uptake of lipids by adipocytes. Similarly, studies in the endothelial communication between adipose ECs and adjacent adipocytes through paracrine signals have suggested that the formation of capillary structures and AT expansion, involves the production of pro-angiogenic factors by adipocytes and ECs, particularly VEGF, and the activation of VEGFR1/VEGFR2 in ECs, as well as the participation of perlecan and endocan. Of note, overexpression of VEGF in AT leading to an increase in blood vessel number and size has been reported. With respect to the immunolocalization of TLR4, FN and TSP-1 around mature adipocytes and in the ECs of the small blood vessels, several studies have showed that ECs and adipocytes express TLR4 on their surface and that activation of this glycoprotein by some of their endogenous ligands such as fibrinogen, FN, HSPGs including perlecan, agrin, and GAL-3 contribute not only to the formation of microvasculature and inflammation, but also to adipocyte differentiation (adipogenesis) and innate immunity. Therefore, it is plausible that perlecan, agrin, CD44, endocan, TLR4, and FN, could be mediating the adipocyte-EC crosstalk, contributing to the growth, expansion and maintenance of AT and the formation of small blood vessels in angiolipoma, which are cellular processes that could be accompanied by the production of VEGF and activation of VEGFR2.
Immunofluorescence also revealed that integrin β1-subunit was localized around the mature adipocytes that were in close proximity to small blood vessels, as well as in the ECs lining the lumen of these vessels and some immune cells. Consistent with this, studies on adipogenesis and integrins have proposed that regulation of the interaction between integrins, particularly integrin α5β1 and αvβ1, and FN contribute to reorganization of the actin cytoskeleton and accomplishment of adipocyte differentiation (adipogenesis). Similarly, studies in the AT vasculature have suggested that the interactions between adipocytes and ECs involving FN, collagens and integrin β1 in the presence of VEGF, could be contributing to the angiogenesis expansion observed in AT. Thus, it is likely that integrin β1 subunit could be mediating the adipocyte-EC crosstalk and contributing to the adipocyte growth, expansion and vascular remodeling in angiolipoma tissues.
Regarding the presence of PECAM-1, endoglin, VE-cadherin and VEGFR2 in the ECs lining the lumen of the tortuous and dilated small blood vessels, it is well known that these molecules together particular proteoglycans such as perlecan, CD44 and endocan and ECM components including the glycoproteins FN, TSP-1 and integrin β1, in combination with certain angiogenic factors produced by activated ECs including VEGF, are key for angiogenesis and vascular remodeling, as well as the regulation of vessel permeability. Of note, modulation of angiogenesis by perlecan through interaction of heparan sulfate side chains with growth factors especially VEGF, and several extracellular matrix components such as FN, TSP-1 and integrin β1, has been suggested. Moreover, interaction fibrinogen-EC involving endothelial integrins such as α1β5 and αvβ3 and non-integrin receptors including PECAM-1, endoglin, VE-cadherin, as well as TLR4 and some of its endogenous ligands including fibrinogen/fibrin, facilitating the EC-EC and IC-EC interactions and, in turn, contributing to small blood vessels formation in angiolipoma, has recently been proposed. Thus, the presence of PECAM-1, endoglin, VE-cadherin VEGFR2, together activated TLR4 in the luminal ECs could be contributing to the AT angiogenesis and in turn, to the regulation of growth, expansion and maintenance of AT in angiolipoma.
In this study, immunostaining also revealed that NRP1 was present in the luminal ECs of the small blood vessels. Consistent with this, several lines of evidence indicate that NRP-1 act as co-receptor for VEGFR2 enabling the binding of VEGF and promoting the angiogenesis. Along these lines, overexpression of NRP1 resulting in the formation of excess capillaries, has been suggested. Additionally, NRP-1 multiprotein complexes formation with VEGFR2, endoglin, HSPGs, endocan, FN, integrin β1 and GAL-1 on the EC surface regulating vascular permeability, has also been suggested. Moreover, recent studies have suggested the colocalization of NRP1 with VE-cadherin and HSPGs, transforming growth factor-β receptors (TGFβRs) regulating adherens junction plasticity and inflammatory response. Therefore, we believe that interaction of NRP1 with the components of adheren junctions and endothelial surface including VE-cadherin, PECAM-1, VEGFR2, endoglin, HSPGs, integrin β1, and GAL-1, could be modulating angiogenesis in angiolipoma.
Interestingly, immunofluorescence also evidenced immunoreactivities for GAL-1, -3 and -8 in angiolipoma tissues on the surface of mature adipocytes that were in close proximity to tortuous and dilated small blood vessels, as well as in the ECs lining the lumen of these vessels. Noteworthy, studies in vitro and in vivo in AT have shown that GAL-1 and GAL-3 are expressed in adipocytes from both humans and animals, being considered as important for AT homeostasis and involved in the modulation of the differentiation and proliferation of adipocytes. Similarly, studies on galectins have suggested that secreted galectins, particularly GAL-1, -3 and -8, can remain in the vicinity of the cell either through binding and cross-linking to the N- and/or O-linked glycan residues of the glycoproteins or GAG side chains of the proteoglycans that are present on the cell surface or in the extracellular milieu. Same studies and others suggest that extracellular galectins can appear arranged into non-covalent bound dimers and oligomeric forms or appear organized in supramolecular multivalent complexes (GAL-glycan lattices) on the cell surface and in the extracellular milieu modulating, through specific signaling pathways, cellular processes including cell-cell communication (crosstalk), cell-matrix adhesion, proliferation, growth, differentiation, migration, and inflammation. It is therefore possible that in angiolipoma GAL-1, GAL-8 and GAL-3 can bind and cross-link the proteoglycans perlecan, agrin, CD44 and endocan and the glycoproteins FN, TSP-1, TLR4 and integrin β1 generating GAL-glycan lattices on the surface of adipocytes and ECs.
Regarding the presence of GAL-1, -3 and -8 in the ECs lining the lumen of small blood vessels, in vitro and in vivo studies have shown that EC activation upregulates the synthesis of GAL-1, -3 and -8, and that they are able to generate Gal-glycan lattices on the surface of ECs mediating the angiogenesis process through interactions with cell-surface glycoproteins that includes PECAM-1 (CD31), VE-cadherin, endoglin (CD105), integrin β1, and VEGFR2, proteoglycans such as perlecan, CD44 and endocan and certain glycoproteins that includes FN and TSP-1. Indeed, studies of GALs and angiogenesis indicate that GAL-1, -3 and -8 promote angiogenesis via activation of VEGFR2 signaling pathway, and that both GAL-3 and GAL-8 interact with integrin β1 subunit regulating the adhesion to extracellular matrix components. Also, formation of GAL-1, NRP-1 and VEGFR2 complexes driving angiogenesis and inducing vascular permeability has been suggested. Other studies report that interaction of GAL-3 with PECAM-1 (CD31), endoglin (CD105) and CD44 stimulates EC migration and angiogenesis. Likewise, binding of GAL-3 to the β-galactoside residues present in the TLR2 and TLR4 ectodomains regulating innate and adaptive responses and promoting angiogenesis, has been proposed. Ultimately, seems likely that in angiolipoma, GAL-1, -3 and -8 acting in concert with their binding partners, form a supramolecular multivalent structure on the surface of mature adipocytes and ECs that might be facilitating the bidirectional communication (crosstalk) between adipocytes and ECs and thus contributing to the mature adipocytic expansion and the new vasculature formation (angiogenesis) or microvasculature remodeling.
Conclusions
We propose that in angiolipoma, GAL-1, -3 and -8 acting in concert with their binding partners form a supramolecular structure on the surface of mature adipocytes and ECs that might be facilitating the crosstalk between adipocytes and ECs, contributing to the mature adipocytic expansion and the new vasculature formation (angiogenesis) or microvasculature remodeling.
Statements and Declarations
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding statement
This work was supported by grant N°2023PGP61 from Fondo Nacional de Ciencia, Tecnología e innovación (FONACIT).
Acknowledgements
The authors thank Biba Arciniegas-Mata for English-editing of this manuscript.
Author contributions
E.A. analyzed the data and wrote the paper; A.D., H.R., A.S. and R.R. analyzed the data and constructed the images; A.D. constructs the scheme; J.P., A.U. and A.M.S examined the biopsies. All authors read and approved the final manuscript.
References
- Alonso S, Rodríguez-Peralto JL, Pérez-Espejo G. Metastasis of cutaneous malignant melanoma to angiolipoma: the tumor-to-tumor metastasis phenomenon. J Cutan Pathol. 2003; 30(5):323-325.
- Lapidoth M, Ben Amitai D, Feinmesser M, Akerman L. Capillary malformation associated with angiolipoma: analysis of 127 consecutive clinic patients. Am J Clin Dermatol. 2008; 9(6):389-392.
- Heng W, Lu L, Wang J. Cellular angiolipoma: a clinicopathological and immunohistochemical study of 12 cases. Am J Dermatopathol. 2013; 35(2):220-225.
- Saggini A, Santonja C, Nájera L, Palmedo G, Kutzner H. Frequent activating PIK3CA mutations in sporadic angiolipoma. J Cutan Pathol. 2021; 48(2):211-216.
- Díaz-Flores L, Gutiérrez R, Pino García M et al. Intussusceptive angiogenesis facilitated by microthrombosis has an important example in angiolipoma. An ultrastructural and immunohistochemical study. Histol Histopathol. 2023; 38(1):29-46.
- Kransdorf MJ, Larsen BT, Goulding, KA, Cumsky JL, Hwang S, Long J. Angiolipoma: a review of 778 lesions in 344 patients. Skeletal Radiol. 2023; 52(3):541-552.
- Arciniegas E, Rojas H, Pineda J, et al. Endothelial TLR4 Activation by endogenous ligands contributes to small blood vessels formation in angiolipoma. J Dermatol Res. 2024; 5(3):1-16. doi.org/10.46889/JDR.2024.5311.
- Dvorak HF, Nagy JA, Feng D, Brown LF, Dvorak AM. Vascular permeability factor/vascular endothelial growth factor and the significance of microvascular hyperpermeability in angiogenesis. Curr Top Microbiol Immunol. 1999; 237:97–132.
- Wu M-H, Ying N-W, Hong T-M, Chiang W-F, Lin Y-T, Chen Y-L. Galectin-1 induces vascular permeability through the neuropilin-1/vascular endothelial growth factor receptor-1 complex. Angiogenesis. 2014. doi.org/10.1007/s10456-014-9431-8.
- Rupnick MA, Panigrahy D, Zhang Ch-Y, et al. Adipose tissue mass can be regulated through the vasculature. PNAS. 2002; 99(16);10730-10735.
- Hausman GJ, Richardson RL. Adipose tissue angiogenesis. J. Anim. Sci. 2004; 82:925–934.
- Alvarez-Llamas G, Szalowska E, de Vries MP, Weening D, Landman K, Hoek A. Characterization of the human visceral adipose tissue secretome. Mol Cell Proteomics. 2007; 6(4):589-600.
- Cao Y. Adipose tissue angiogenesis as a therapeutic target for obesity and metabolic diseases. Nat Rev Drugs Discov. 2010; 9:107-115.
- Christiaens V, Lijnen HR. Angiogenesis and development of adipose tissue. Mol Cell Endocrinol. 2010; 318:2–9.
- Crewe C, An YA, Scherer PE. The ominous triad of adipose tissue dysfunction: inflammation, fibrosis, and impaired angiogenesis. J Clin Invest. 2017; 127(1):74-82.
- Ruiz-Ojeda FJ, Méndez-Gutiérrez A, Aguilera MC, Plaza-Díaz J. Extracellular matrix remodeling of adipose tissue in obesity and metabolic diseases. Int J Mol Sci. 2019, 20, 4888. doi.org/10.3390/ijms20194888.
- Herold J, Kalucka J. Angiogenesis in adipose tissue: The interplay between adipose and endothelial cells. Front Physiol. 2021; 11;624903. doi.org/10.3389/fphys.2020.624903.
- Sun K, Li X, Scherer PE. Extracellular matrix (ECM) and fibrosis in adipose tissue: overview and perspectives. Compr Physiol. 2023; 13(1):4387–4407. doi.org/10.1002/cphy.c220020.
- Corvera S. Cellular Heterogeneity in Adipose Tissues. Annu Rev Physiol. 2021; 10; 83:257–278.
- Corvera S, Solivan-Rivera J, Loureiro ZY. Angiogenesis in adipose tissue and obesity. Angiogenesis. 2022; 25:439–453.
- Maniyadath B, Zhang Q, Gupta RK. Mandrup S. Adipose tissue at single cell resolution. Cell Metab. 2023; 35(3):386–413.
- Santillana N, Astudillo-Guerrero C, D’Espessailles A, Cruz G. White adipose tissue dysfunction: pathophysiology and emergent measurements. Nutrients. 2023; 15, 1722.
- Auger Ch, Kajimura S. Adipose tissue remodeling in pathophysiology. Annu Rev Pathol. 2023; 18: 71–93.
- Johnston EK, Abbott RD. Adipose tissue development relies on coordinated extracellular matrix remodeling, angiogenesis, and adipogenesis. Biomedicines. 2022; 10, 2227. doi.org/10.3390/biomedicines10092227.
- Iacobini C, Vitale M, Haxhi J, Menini S, Pugliese G. Impaired remodeling of white adipose tissue in obesity and aging: from defective adipogenesis to adipose organ dysfunction. Cells. 2024; 13, 763. doi.org/10.3390/cells13090763.
- Arderiu G, Civit-Urgell A, Díez-Caballero A , Moscatiello F, Ballesta C, Badimon L. Differentiation of adipose tissue mesenchymal stem cells into endothelial cells depends on fat depot conditions: regulation by miRNA. Cells. 2024; 13, 513. doi.org/10.3390/cells13060513.
- Mo Y-Y, Han Y-X, Xu S-N, et al. Adipose tissue plasticity: a comprehensive definition and multidimensional insight. Biomolecules. 2024; 14, 1223. doi.org/10.3390/biom14101223.
- Chaurasiya V, Nidhina Haridas PA, Olkkonen VM. Adipocyte-endothelial cell interplay in adipose tissue physiology. Biochem Pharmacol. 2024; 222, 116081. doi.org/10.1016/j.bcp.2024.116081.
- Aoki S, Toda S, Sakemi T, Sugihara H. Coculture of endothelial cells and mature adipocytes actively promotes immature preadipocyte development in vitro. Cell Struct Function. 2003; 28:55-60.
- Rojas-Rodriguez R, Gealekman O, Kruse ME, et al. Adipose tissue angiogenesis assay. Methods Enzymol. 2014; 537: 75–91.
- Blázquez Medela AM, Penton A, Bostrom KI, Saparov A, Jumabay M. Generation of Vascular Networks from Adipocytes in vitro. Int J Cell Sci Mol Biol. 2019; 6(2):1-12.
- Sabaratnam R, Svenningsen P. Adipocyte-endothelium crosstalk in obesity. Front Endocrinol. 2021; 12;681290. doi.org/10,3389/fendo.2021,681290.