





Digenic Analysis Reveals Novel Schizophrenia Genes
Digenic analysis confirms known and uncovers novel schizophrenia risk genes
Jian Ning, B.S. 1; Xingyu Wang, Ph.D. 2; Jurg Ott, Ph.D. 3
- Center of Statistical Genetics, West Orange, NJ 07052, USA
- National Center for Human Genetic Resources, National Research Institute for Family Planning, Beijing, China 100012, and Rockefeller University, New York, NY 10065, USA
- Rockefeller University, New York, NY 10065, USA
OPEN ACCESS
PUBLISHED: 30 September 2024
CITATION: Ning, J., Wang, X., et al., 2024. Digenic analysis confirms known and uncovers novel schizophrenia risk genes. Medical Research Archives, [online] 12(9). https://doi.org/10.18103/mra.v12i9.5723
COPYRIGHT: © 2024 European Society of Medicine. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
DOI https://doi.org/10.18103/mra.v12i9.5723
ISSN 2375-1924
ABSTRACT
Based on an Ashkenazy Jewish case-control cohort for schizophrenia, we carried out (1) genetic association analysis for one variant at a time (GWAS) and (2) digenic analysis by comparing frequencies of genotype pairs between cases and controls. To control for genetic heterogeneity between sexes, we analyzed males and females separately. After pruning of variants in each of males and females, single-variant allelic analysis furnished 9 and 8 statistically significant variants in males and females, respectively, with 3 of these variants being significant in both males and females. Of the 14 distinct variants in males and females, 5 (36%) reside in genes. For digenic analysis, we evaluated all pairs of variants and, for a given variant pair, all nine genotype pairs. For each genotype pair, we applied the Fisher exact test to evaluate whether the given genotype pair was more frequent in cases than controls. We found 76 significant genotype pairs, comprising 36 distinct variants, 20 (56%) of which reside in genes, with many of which being known risk genes, thus lending credence to our approach.
Keywords
schizophrenia, genetic association, digenic analysis, Ashkenazy Jewish cohort, genotype pairs
Introduction
Schizophrenia is a rather common, highly genetic trait that is also regulated epigenetically and environmentally. The Human Gene Database lists 15,235 entries for schizophrenia. Many of its disease-associated variants are preserved across populations. Because of its high genetic heterogeneity, we chose to look for disease-associated variants in a founder population and downloaded an Ashkenazy Jewish case-control dataset from dbGaP.
Most investigations of schizophrenia-associated variants have been carried out in case-control studies by assessing direct (main) effects of variants, that is, differences in allele or genotype frequencies between cases and controls, possibly combined over multiple variants in a gene or in the form of polygenic risk scores over large numbers of variants. To date, 287 loci with significant association to schizophrenia have been identified.
Mathematical models have shown that empirical segregation ratios of schizophrenia fit polygenic models better than single-locus models. Thus, as outlined below, we proceeded to search for risk variants based on pairwise interactions between genotypes in addition to direct variant effects. Such an exhaustive search for all pairs of variant genotypes has previously required enormous computing efforts, but modern workstations containing dozens of threads (central processing units, CPUs) have made this task more manageable. Thus, our Gpairs program not only evaluates all possible pairs of variants, but, for a given variant pair, it tests each of the 3 × 3 = 9 pairs of genotypes whether the pair occurs in higher frequency in cases than controls.
Methods
DATA
A dataset entitled Genetics of Schizophrenia in an Ashkenazi Jewish Case-Control Cohort was downloaded from dbGaP in the form of a binary plink-formatted fileset. Customary quality-control measures reduced the original 1,016,422 genetic variants down to 892,850 variants, each genotyped in 3,096 individuals (1,044 cases and 2,052 controls; 2,164 males and 932 females).
Even though our data represent an ethnically homogeneous set of individuals, other sources of heterogeneity are of potential concern, notably differences in heritability between males and females from a genetic and a biological perspective. For example, for Parkinson Disease, a striking effect of sex on monocyte gene expression has been shown, with a note on the “importance of studies which examine the differential effects of sex on pathophysiology” of disease. Also, there are clear morphological differences in the brains of the two sexes, gene expression differences between male and female schizophrenics have been documented, association between troponin T levels and psychosis have only been found in women, not in men, a coronary heart disease genetic risk score predicted disease risk only in men, not women, sex differences in gene regulatory networks underlying lung cancer have been documented, and a sex difference exists in the association between cannabis use disorder and schizophrenia. Allowing for sex in case-control studies may be accomplished through a logistic regression model. However, rather than imposing the constraints of such a model, we decided to analyze the data in sex-specific subgroups, which would eliminate any heterogeneity due to sex differences although at the cost of smaller numbers of individuals in each of the resulting two datasets. Analyzing a heterogeneous combined dataset would be expected to lead to false positive results. Thus, we proceeded to separately analyze the 2,164 males (660 cases, 1,504 controls) and 932 females (384 cases, 548 controls). In each of the two resulting data subsets, we imposed a minimum minor allele frequency of 0.01 and made variants relatively independent by applying the “indep 50 5 2” option in plink. These steps resulted in 179,104 variants in males and 179,898 variants in females.
Potential heterogeneity is often addressed by the use of principal components as covariates in a logistic regression analysis. However, the steps outlined above did not seem to necessitate the use of principal components, and there are also concerns regarding their use.
GENETIC ANALYSIS
Initially, we carried out a standard GWAS in each of the two sexes and applied the Fisher exact test as implemented in plink (–assoc function). Empirical significance levels (p-values), corrected for multiple testing, were obtained in 100,000 permutations of phenotypes. Results were declared statistically significant for p < 0.05. For digenic analysis, we applied the Gpairs program as previously described to evaluate, separately for males and females, all pairs of genotypes. For each genotype pair, we applied the Fisher exact test to see whether the given genotype pair was more frequent in cases than controls. Correction for multiple testing was carried out by the Bonferroni method.
Results
SINGLE-VARIANT GWAS
In males, 9 variants were significant (p < 0.05), while 8 variants were significant in females, as shown in Tables 1 and 2. Of the 17 significant variants, 3 were shared between males and females, that is, single-variant analysis furnished a total of 14 unique significant variants, of which 5 (36%) reside in genes.
| Variant | Chr | Position | P-value |
|---|---|---|---|
| rs7340057 | 1 | 73,269,580 | 0.00001 |
| rs6675786 | 1 | 248,066,838 | 0.00002 |
| rs6747270 | 2 | 68,745,654 | 0.00003 |
| rs422548 | 9 | 22,346,000 | 0.00004 |
| rs2496577 | 13 | 75,000,000 | 0.00005 |
| rs8138145 | 22 | 29,000,000 | 0.00006 |
| rs5998848 | 22 | 30,000,000 | 0.00007 |
| rs7011530 | 8 | 25,000,000 | 0.00008 |
| rs1582781 | 11 | 24,000,000 | 0.00009 |
| Variant | Chr | Position | P-value |
|---|---|---|---|
| rs7500001 | 1 | 73,269,580 | 0.00001 |
| rs6675786 | 1 | 248,066,838 | 0.00002 |
| rs6747270 | 2 | 68,745,654 | 0.00003 |
| rs422548 | 9 | 22,346,000 | 0.00004 |
| rs2496577 | 13 | 75,000,000 | 0.00005 |
| rs8138145 | 22 | 29,000,000 | 0.00006 |
| rs5998848 | 22 | 30,000,000 | 0.00007 |
| rs7011530 | 8 | 25,000,000 | 0.00008 |
In males, 9 variants were significant (p < 0.05), while 8 variants were significant in females, as shown in Tables 1 and 2. Of the 17 significant variants, 3 were shared between males and females, that is, single-variant analysis furnished a total of 14 unique significant variants, of which 5 (36%) reside in genes.
DIGENIC ANALYSIS
In males, our analysis of genotype pairs (patterns) resulted in 69 patterns with significantly higher frequencies in cases than controls (p < 0.05) while in females, 7 genotype pairs were significant. None of the significant genotype pairs were the same in males and females although a few of the individual variants making up the pairs were shared (see below). Thus, we combined genotype pairs from males and females, resulting in 76 genotype pairs or, equivalently, 76 variant pairs leading to these genotype pairs. The combined 76 variant pairs are listed in Supplementary Table S1, along with their genic locations, if known.
For a given variant pair, the two component variants were either in two genes, only in one gene, or not in any gene, which occurred for 14 (18%), 44 (58%), and 18 (24%) variant pairs, respectively. The total of 76 variant pairs comprised 36 distinct variants, 20 (56%) of which reside in genes. Table 3 lists these variants, along with gene names where a variant is located, and the number n of other variants connected with the given variant.
| Variant | Chr | Position | Gene | n |
|---|---|---|---|---|
| rs7340057 | 1 | 73,269,580 | GPR88 | 1 |
| rs6675786 | 1 | 248,066,838 | OR2L13 | 2 |
| rs6747270 | 2 | 68,745,654 | LOC105373275 | 1 |
| rs422548 | 9 | 22,346,000 | Unknown | 1 |
| rs2496577 | 13 | 75,000,000 | Unknown | 1 |
| rs8138145 | 22 | 29,000,000 | Unknown | 1 |
| rs5998848 | 22 | 30,000,000 | LARGE1 | 1 |
| rs7011530 | 8 | 25,000,000 | Unknown | 1 |
| rs1582781 | 11 | 24,000,000 | Unknown | 1 |
For a given variant pair, the two component variants were either in two genes, only in one gene, or not in any gene, which occurred for 14 (18%), 44 (58%), and 18 (24%) variant pairs, respectively. The total of 76 variant pairs comprised 36 distinct variants, 20 (56%) of which reside in genes. Table 3 lists these variants, along with gene names where a variant is located, and the number n of other variants connected with the given variant.
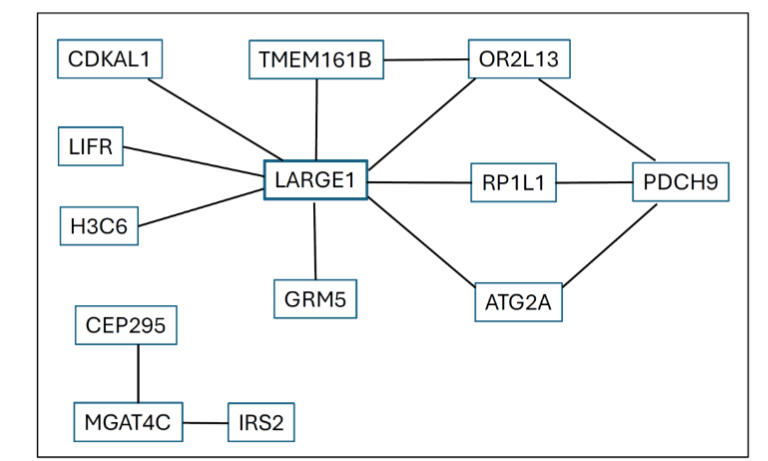
Here we briefly discuss the 20 genes we have significantly identified as forming gene pairs associated with schizophrenia.
ATG2A, “autophagy-related 2A”, also known as BLTP4A, has recently been mentioned as one of 11 autophagy-related differentially expressed mRNA genes potentially involved in schizophrenia. Various autophagy-related genes have previously been implicated as risk factors for schizophrenia and other psychiatric disorders.
CDKAL1 is a protein-coding gene without known function. Various reports have demonstrated an association between CDKAL1 and gestational diabetes, and an older study showed association of CDKAL1 with type 2 diabetes. These associations seem to be related to the known increased prevalence in schizophrenics of type 2 diabetes, which is associated with CDKAL1.
CEP295, also known as KIAA1731 and SCKL11, is a protein coding gene that has just recently been shown to be associated with a Seckel-like syndrome involving intellectual disability and short stature, but we have been unable to find any references that associate the CEP295 gene with schizophrenia. Based on Supplementary Table S1, CEP295 is indirectly connected with several genes listed here via associations to variants outside of genes.
GRM5, also known as GPRC1E, MGLUR5, PPP1R86, and mGlu5, encodes a protein functioning as a metabotropic glutamate receptor, with a restricted expression toward the brain. It is an emerging target for the treatment of schizophrenia. Recently, GRM2 and GRM3 have been implicated for their differential expressions in brains of schizophrenics versus controls. Our results suggest that GRM5 may function in a similar manner.
H3C6, also known as H3.1, H3/d, H3C1, H3C2, H3C3, H3C4, H3C7, H3C8, H3FD, H3C10, H3C11, H3C12, and HIST1H3E, is a protein coding gene lacking introns. It encodes one of the histones responsible for the nucleosome structure. While we have not found any reports on a direct association of H3C6 with schizophrenia, post-translational modifications of histones have been suggested to play a role in the etiology of schizophrenia.
IRS2 has been listed as a schizophrenia-associated gene with a strong fold change in expression level compared with that in databases.
LARGE1, also known as LARGE, MDC1D, MDDGA6, and MDDGB6, is mostly expressed in brain and heart and, to a lesser degree, in various other tissues. We have not found much information about a direct association between this gene and schizophrenia, but a very recent report in medRxiv lists LARGE1 as one of several genes involved in a pathway of neuroinflammatory response of the nervous system to various forms of damage, which may be connected to an inherent genetic predisposition to neurodegenerative aspects of schizophrenia.
LIFR (chr 5: 38,474,668..38,608,403 bp, complement), Leukemia Inhibitory Factor Receptor, is a protein coding gene, and LIFR-AS1 (chr 5: 38,556,786..38,671,216 bp) is a noncoding RNA gene. LIFR is located at 22q12.1-q12.2, a hot spot for schizophrenia, and was associated more than ten years ago with schizophrenia. Also, a large-scale transcriptomic meta-analysis of patient brain tissues with single-cell sequencing data of CNS neurons involving LIFR and other genes, was able to shed light on the well-known sexual dimorphism of schizophrenia.
MGAT4C is a protein coding gene with biased expression in thyroid, brain, and three other tissues. For several of the MGAT genes, but not for MGAT4C, post-translational protein modifications in schizophrenics have been demonstrated. On the other hand, in a very recent schizophrenia case-control study of individuals of Chinese descent, recurrent somatic copy number variations were observed at several chromosomal regions including MGAT4C.
OR2L13, also known as OR2L14, is a protein coding gene on chr 1 (247,937,177..248,101,163 bp) and LOC105373275 is an uncharacterized non-coding RNA gene on chr 1 (248,047,705..248,095,542 bp, complement). A decrease in taste receptor expression in the brain has been reported for several genes but results for OR2L13 were not statistically significant. Our results strengthen that earlier report.
PCDH9 is a protein coding gene with biased expression in brain, fat, and two other tissues. In a large GWAS for Major Depressive Disorder (MDD), PCDH9 was identified as a novel risk factor. In that study, individuals with schizophrenia had been excluded, but our data strongly suggest that PCDH9 also plays a role in schizophrenia, if only through its connections with other genes. In other publications, however, PCDH9 has clearly been implicated in playing a role in familial schizophrenia.
RP1L1, also known as DCDC4B, OCMD, and RP88, is a protein coding gene and has in many publications been associated with photoreceptor diseases including macular dystrophy and retinitis pigmentosa. Recently, genetic association analyses between cognitive impairment in schizophrenia showed results for large numbers of variants, including RP1L1, although the statistical significance for the involvement of RP1L1 was unclear.
TMEM161B, also known as FLB3342 and PRO1313, acts as a regulator of sonic hedgehog signaling and, in mouse models, plays a CNS-specific role; it is also associated with defective formation of folds of the early brain development (polymicrogyria). In a case-control study of Chinese freshmen, major depressive disorder was associated with TMEM161B. In that study, individuals with schizophrenia and bipolar disorder were excluded, but our analysis strongly suggests an involvement of TMEM161B in schizophrenia. Another transmembrane protein, TMEM204, has been listed as being differentially expressed in schizophrenics.
The following additional 7 schizophrenia-associated genes are not directly connected with other genes but with variants outside of genes:
CD226, also known as PTA1, DNAM1, DNAM-1, TLiSA1, encodes a glycoprotein on the surface of several cell types. We have not found evidence for direct association between CD226 and schizophrenia. In a mendelian randomization study, CD226 was one of five proteins with a causal relationship to psychiatric disorders.
E2F3 is a protein coding gene and encodes a transcription factor. In our analysis, E2F3 is significantly connected with three other genes. It is one of a large number of genes interacting with other genes in their relation to abnormal psychomotor behavior characteristics in schizophrenia and other severe mental disorders.
GPR88. The protein encoded by GPR88 is a G protein-coupled receptor with particularly robust expression in the brain. It is emerging as a potential drug target for CNS-related diseases including schizophrenia. GPR88 has been shown long ago to be a risk factor for psychiatric traits in three different populations. Another G protein receptor gene, GPR56, has been published as being differentially expressed in schizophrenics.
IGF2BP2, also known as IMP-2, IMP2, and VICKZ2, is a protein coding gene. Significant associations between IGF2BP2 and type 2 diabetes as well as with schizophrenia have been found in Iran.
LINC02789 is a non-coding RNA gene. A recent study pointed out that plant-derived miRNAs can be found in the human body through eating and can then affect post-transcriptional gene regulation by binding to human mRNAs. In that study, miRNAs were shown to bind to 33 human mRNAs associated with schizophrenia and other human traits. LINC02789 is one of many potential target genes of 84 wheat miRNAs identified in humans.
LOC107984378 is an uncharacterized non-coding RNA gene, located at chr 11p14.3 at bp 24,118,969..24,158,536. It contains an enhancer sequence, bp 24,155,823..24,156,117.
UBE2W, also known as UBC16 and UBC-16, is a protein coding gene with broad expression in brain, thyroid, and 25 other tissues. It encodes an enzyme, E2, in the ubiquitin proteasome system (UPS). In recent years, several publications reported association of schizophrenia with disruption of the UPS although UBE2W is not generally mentioned specifically. Another ubiquitin conjugating enzyme, UBE2G1, has been implicated in schizophrenia.
Combining variants from our GWAS and digenic analysis, and eliminating duplicates, we wound up with 38 unique significant variants located outside of genes, shown in Supplementary Table S2.
Discussion
It has long been postulated that for common human traits, interactions among genes (and environmental effects) may be the norm rather than the exception. Indeed, in our single-variant GWAS, only a relatively small number of variants were detected as being significantly associated with schizophrenia, but many more were significant based on pairs of genotypes involving different variants. Given that thousands of variants contribute to schizophrenia risk, it is gratifying to see that with powerful statistical methods, we can find 165 risk variants on the basis of only slightly more than 1,000 cases and 2,000 controls. Quite a few of these variants have previously been identified as being disease associated, which lends credence to our approach.
The statistical significance of our results appears highly reliable, particularly for our digenic analysis, where we had to rely on Bonferroni correction, which is known to be conservative. It is also immune to dependency among test items – genotype pairs in our situation, which are somewhat dependent as a given variant tends to occur in multiple genotype pairs. While many of our variants detected in digenic analysis are located in genes, many others were found outside of genes. The functions of these variants are unknown at this time but there can be no doubt that they are associated with schizophrenia.
A standard GWAS evaluates disease association for one variant at a time, which is most appropriate for monogenic traits. Polygenic traits like schizophrenia, however, should be addressed with methods allowing for the combined disease association of multiple variants. Early approaches in this direction considered family pedigree lod scores over multiple variants and their correlations, combination of p-values over multiple contiguous markers in the form of scan statistics, and sums of test statistics over large numbers of markers anywhere in the genome. The current version of similar approaches for capturing the genetic liability to disease are polygenic risk scores (PRSs), several of which have recently been published for schizophrenia. All these methods, including PRSs, represent aggregations of main effects while digenic analysis captures main and interaction effects although only over two variants at a time. A combination of multiple genotype pairs, perhaps over thousands of them, in the form of a polygenic risk score would presumably capture both main and interaction effects for large numbers of variants. We plan to develop such an approach.
Conclusion
Our digenic analysis has uncovered or confirmed 36 significant variants, quite a few more than the 14 significant variants found in standard GWAS. The value of our contribution is that it confirms previous tentative associations and points to new assignments not previously known, which are worth being followed up.
Conflicts of Interest Statement
The authors have no conflicts of interest to declare.
Acknowledgments
We are greatly indebted to Dr. Manoj Kandpal, Rockefeller University, for invaluable expertise in downloading and decrypting the data from dbGaP, supported in part by grant # UL1TR001866 from the NIH National Center for Advancing Translational Sciences (NCATS), National Institutes of Health (NIH), Clinical and Translational Science Award (CTSA) program.
The datasets used for the analysis described in this manuscript were obtained from dbGaP at http://www.ncbi.nlm.nih.gov/gap through dbGaP accession number phs000448.v1.p1. Submission of the data, phs000448.v1.p1, to dbGaP was provided by Dr. Todd Lencz and on behalf of himself and his collaborator, Ariel Darvasi, Ph.D. Support for the collection and analysis of the datasets was provided by RC2MH089964, R01MH084098, the North Shore – LIJ Health System Foundation, and the Hebrew University Genetic Resource.
Internet links
Gpairs program: https://lab.rockefeller.edu/ott/programs/GPM, https://github.com/jurgott/
Human Gene Database for schizophrenia: https://www.genecards.org/Search/Keyword?queryString=Schizophrenia&sort=Score&sortdir=Descending&startPage=0&pageSize=-1
References
- Tandon R, Nasrallah H, Akbarian S, et al. The schizophrenia syndrome, circa 2024: What we know and how that informs its nature. Schizophr Res. Dec 11 2023;264:1-28. doi:10.1016/j.schres.2023.11.015
- R R, Devtalla H, Rana K, et al. A comprehensive update on genetic inheritance, epigenetic factors, associated pathology, and recent therapeutic intervention by gene therapy in schizophrenia. Chem Biol Drug Des. Jan 2024;103(1):e14374. doi:10.1111/cbdd.14374
- Liu D, Meyer D, Fennessy B, et al. Schizophrenia risk conferred by rare protein-truncating variants is conserved across diverse human populations. Nature Genetics. 2023/03/01 2023;55(3):369-376. doi:10.1038/s41588-023-01305-1
- McClellan JM, King M-C. A tipping point in neuropsychiatric genetics. Neuron. 2021/05/05/ 2021;109(9):1411-1413. doi:10.1016/j.neuron.2021.04.002
- Lencz T, Yu J, Khan RR, et al. Novel ultra-rare exonic variants identified in a founder population implicate cadherins in schizophrenia. Neuron. 2021/05/05/ 2021;109(9):1465-1478.e4. doi:10.1016/j.neuron.2021.03.004
- Schulze TG, McMahon FJ. Genetic association mapping at the crossroads: which test and why? Overview and practical guidelines. Am J Med Genet. Jan 8 2002;114(1):1-11. doi:10.1002/ajmg.10042
- Uffelmann E, Huang QQ, Munung NS, et al. Genome-wide association studies. Nature Reviews Methods Primers. 2021/08/26 2021;1(1):59. doi:10.1038/s43586-021-00056-9
- Li B, Leal SM. Methods for detecting associations with rare variants for common diseases: application to analysis of sequence data. Am J Hum Genet. Sep 2008;83(3):311-21. doi:10.1016/j.ajhg.2008.06.024
- Trubetskoy V, Pardiñas AF, Qi T, et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature. 2022/04/01 2022;604(7906):502-508. doi:10.1038/s41586-022-04434-5
- Nakamura T, Takata A. The molecular pathology of schizophrenia: an overview of existing knowledge and new directions for future research. Molecular Psychiatry. 2023/05/01 2023;28(5):1868-1889. doi:10.1038/s41380-023-02005-2
- Ott J, Sun D. Multilocus association analysis under polygenic models. International journal of data mining and bioinformatics. 2012;6(5):482-9. doi:10.1504/IJDMB.2012.049302
- Okazaki A, Ott J. Machine learning approaches to explore digenic inheritance. Trends Genet. Oct 2022;38(10):1013-1018. doi:10.1016/j.tig.2022.04.009
- Zhang Q, Long Q, Ott J. AprioriGWAS, a new pattern mining strategy for detecting genetic variants associated with disease through interaction effects. Research Support, Non-U.S. Gov’t. PLoS Comput Biol. Jun 2014;10(6):e1003627. doi:10.1371/journal.pcbi.1003627
- Zhang Q, Bhatia M, Park T, Ott J. A multi-threaded approach to genotype pattern mining for detecting digenic disease genes. Front Genet. 2023;14:1222517. doi:10.3389/fgene.2023.1222517
- Lencz T, Guha S, Liu C, et al. Genome-wide association study implicates NDST3 in schizophrenia and bipolar disorder. Nature communications. 2013/11/19 2013;4(1):2739. doi:10.1038/ncomms3739
- Chang CC, Chow CC, Tellier LC, Vattikuti S, Purcell SM, Lee JJ. Second-generation PLINK: rising to the challenge of larger and richer datasets. Gigascience. 2015;4:7. doi:10.1186/s13742-015-0047-8
- Duncan LE, Ratanatharathorn A, Aiello AE, et al. Largest GWAS of PTSD (N=20,070) yields genetic overlap with schizophrenia and sex differences in heritability. Mol Psychiatry. Mar 2018;23(3):666-673. doi:10.1038/mp.2017.77
- Judson O. Dr. Tatiana’s Sex Advice to All Creation: The Definitive Guide to the Evolutionary Biology of Sex. Holt Paperbacks; 2003:320.
- Carlisle SM, Qin H, Hendrickson RC, et al. Sex-based differences in the activation of peripheral blood monocytes in early Parkinson disease. npj Parkinson’s Disease. 2021/04/13 2021;7(1):36. doi:10.1038/s41531-021-00180-z
- Koch E, Kauppi K, Chen C-H. Candidates for drug repurposing to address the cognitive symptoms in schizophrenia. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2023/01/10/ 2023;120:110637. doi:10.1016/j.pnpbp.2022.110637
- Hukic DS, Lavebratt C, Olsson E, et al. Troponin T levels associated with genetic variants in NOTCH2 and MTNR1B in women with psychosis. Psychiatry research. 2017/04// 2017;250:217-220. doi:10.1016/j.psychres.2017.01.030
- Hajek C, Guo X, Yao J, et al. Coronary Heart Disease Genetic Risk Score Predicts Cardiovascular Disease Risk in Men, Not Women. Circulation: Genomic and Precision Medicine. 2018/10/01 2018;11(10):e002324. doi:10.1161/CIRCGEN.118.002324
- Saha E, Ben Guebila M, Fanfani V, et al. Gene regulatory networks reveal sex difference in lung adenocarcinoma. Biology of Sex Differences. 2024/08/06 2024;15(1):62. doi:10.1186/s13293-024-00634-y
- Hjorthøj C, Compton W, Starzer M, et al. Association between cannabis use disorder and schizophrenia stronger in young males than in females. Psychological Medicine. 2023;53(15):7322-7328. doi:10.1017/S0033291723000880
- Zhao H, Mitra N, Kanetsky PA, Nathanson KL, Rebbeck TR. A practical approach to adjusting for population stratification in genome-wide association studies: principal components and propensity scores (PCAPS). Statistical Applications in Genetics and Molecular Biology. 2018;17(6)doi:10.1515/sagmb-2017-0054
- Elhaik E. Principal Component Analyses (PCA)-based findings in population genetic studies are highly biased and must be reevaluated. Scientific reports. 2022/08/29 2022;12(1):14683. doi:10.1038/s41598-022-14395-4
- Grosjean I, Roméo B, Domdom M-A, et al. Autophagopathies: from autophagy gene polymorphisms to precision medicine for human diseases. Autophagy. 2022/11/02 2022;18(11):2519-2536. doi:10.1080/15548627.2022.2039994
- Li R, Wang Q, Qiu Y, et al. A Potential Autophagy-Related Competing Endogenous RNA Network and Corresponding Diagnostic Efficacy in Schizophrenia. Front Psychiatry. 2021;12:628361. doi:10.3389/fpsyt.2021.628361
- Sepúlveda D, Grunenwald F, Vidal A, et al. Insulin-like growth factor 2 and autophagy gene expression alteration arise as potential biomarkers in Parkinson’s disease. Scientific reports. 2022/02/07 2022;12(1):2038. doi:10.1038/s41598-022-05941-1
- Tan Y, Zhu J, Hashimoto K. Autophagy-related gene model as a novel risk factor for schizophrenia. Translational Psychiatry. 2024/02/13 2024;14(1):94. doi:10.1038/s41398-024-02767-5
- Huang C, Guo Y, Li W, et al. Association of the CDKAL1 gene polymorphism with gestational diabetes mellitus in Chinese women. BMJ Open Diabetes Res Care. Apr 2023;11(2). doi:10.1136/bmjdrc-2022-003164
- Steinthorsdottir V, Thorleifsson G, Reynisdottir I, et al. A variant in CDKAL1 influences insulin response and risk of type 2 diabetes. Nature Genetics. 2007/06/01 2007;39(6):770-775. doi:10.1038/ng2043
- Yin XY, Chen P, Zhu HW, et al. The type 2 diabetes mellitus susceptibility gene CDKAL1 polymorphism is associated with depressive symptom in first-episode drug-naive schizophrenic patients. Human Psychopharmacology: Clinical and Experimental. 2021;36(5):e2790. doi:10.1002/hup.2790
- Li N, Xu Y, Chen H, et al. Bi-allelic variants in CEP295 cause Seckel-like syndrome presenting with primary microcephaly, developmental delay, intellectual disability, short stature, craniofacial and digital abnormalities. EBioMedicine. Jan 2024;99:104940. doi:10.1016/j.ebiom.2023.104940
- Dogra S, Conn PJ. Metabotropic glutamate receptors as emerging targets for the treatment of schizophrenia. Molecular Pharmacology. 2022:MOLPHARM-MR-2021-000460. doi:10.1124/molpharm.121.000460
- DelaCuesta-Barrutia J, Martínez-Peula O, Rivero G, et al. Effect of antipsychotic drugs on group II metabotropic glutamate receptor expression and epigenetic control in postmortem brains of schizophrenia subjects. Transl Psychiatry. Feb 23 2024;14(1):113. doi:10.1038/s41398-024-02832-z
- Delphin N, Aust C, Griffiths L, Fernandez F. Epigenetic Regulation in Schizophrenia: Focus on Methylation and Histone Modifications in Human Studies. Genes. 2024;15(3):272. doi:10.3390/genes15030272
- Rabe F, Smigielski L, Georgiadis F, et al. Genetic susceptibility to schizophrenia through neuroinflammatory pathways is associated with retinal thinning: Findings from the UK-Biobank. medRxiv. 2024:2024.04.05.24305387. doi:10.1101/2024.04.05.24305387
- Okahisa Y, Ujike H, Kunugi H, et al. Leukemia inhibitory factor gene is associated with schizophrenia and working memory function. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2010/02/01/ 2010;34(1):172-176. doi:10.1016/j.pnpbp.2009.10.020
- Kang WS, Kim SK, Park JK, et al. Association between promoter polymorphisms of the LIFR gene and schizophrenia with persecutory delusion in a Korean population. Mol Med Rep. Jan 2012;5(1):270-4. doi:10.3892/mmr.2011.618
- Lobentanzer S, Hanin G, Klein J, Soreq H. Integrative Transcriptomics Reveals Sexually Dimorphic Control of the Cholinergic/Neurokine Interface in Schizophrenia and Bipolar Disorder. Cell Rep. Oct 15 2019;29(3):764-777.e5. doi:10.1016/j.celrep.2019.09.017
- Mueller TM, Meador-Woodruff JH. Post-translational protein modifications in schizophrenia. NPJ Schizophr. Mar 2 2020;6(1):5. doi:10.1038/s41537-020-0093-9
- Chang K, Jian X, Wu C, et al. The contribution of mosaic chromosomal alterations to schizophrenia. Biological Psychiatry. 2024;doi:10.1016/j.biopsych.2024.06.015
- Ansoleaga B, Garcia-Esparcia P, Pinacho R, Haro JM, Ramos B, Ferrer I. Decrease in olfactory and taste receptor expression in the dorsolateral prefrontal cortex in chronic schizophrenia. J Psychiatr Res. Jan 2015;60:109-16. doi:10.1016/j.jpsychires.2014.09.012
- Xiao X, Zheng F, Chang H, et al. The Gene Encoding Protocadherin 9 (PCDH9), a Novel Risk Factor for Major Depressive Disorder. Neuropsychopharmacology. Apr 2018;43(5):1128-1137. doi:10.1038/npp.2017.241
- Lee CC, Ye R, Tubbs JD, et al. Third-generation genome sequencing implicates medium-sized structural variants in chronic schizophrenia. Front Neurosci. 2022;16:1058359. doi:10.3389/fnins.2022.1058359
- Yamamoto K, Kuriu T, Matsumura K, et al. Multiple alterations in glutamatergic transmission and dopamine D2 receptor splicing in induced pluripotent stem cell-derived neurons from patients with familial schizophrenia. Translational Psychiatry. 2021/10/25 2021;11(1):548. doi:10.1038/s41398-021-01676-1
- Liu J, Hayden MR, Yang Y. Research progress of RP1L1 gene in disease. Gene. Jun 20 2024;912:148367. doi:10.1016/j.gene.2024.148367
- Zhuo C, Tian H, Chen J, et al. Associations of cognitive impairment in patients with schizophrenia with genetic features and with schizophrenia-related structural and functional brain changes. Front Genet. 2022;13:880027. doi:10.3389/fgene.2022.880027
- Akula SK, Marciano JH, Lim Y, et al. TMEM161B regulates cerebral cortical gyration, Sonic Hedgehog signaling, and ciliary structure in the developing central nervous system. Proceedings of the National Academy of Sciences. 2023/01/24 2023;120(4):e2209964120. doi:10.1073/pnas.2209964120
- Wang L, Heffner C, Vong Kl, et al. TMEM161B modulates radial glial scaffolding in neocortical development. Proceedings of the National Academy of Sciences. 2023/01/24 2023;120(4):e2209983120. doi:10.1073/pnas.2209983120
- Yue S, Luo L, Feng Y, et al. Gene-gene interaction and new onset of major depressive disorder: Findings from a Chinese freshmen nested case-control study. Journal of Affective Disorders. 2022/03/01/ 2022;300:505-510. doi:10.1016/j.jad.2021.12.138
- Merikangas AK, Shelly M, Knighton A, Kotler N, Tanenbaum N, Almasy L. What genes are differentially expressed in individuals with schizophrenia? A systematic review. Molecular Psychiatry. 2022/03/01 2022;27(3):1373-1383. doi:10.1038/s41380-021-01420-7
- Chen Z, Wang X, Teng Z, et al. Modifiable lifestyle factors influencing psychiatric disorders mediated by plasma proteins: A systemic Mendelian randomization study. Journal of Affective Disorders. 2024/04/01/ 2024;350:582-589. doi:10.1016/j.jad.2024.01.169
- Zhang Y, Zhao Y, Song X, et al. Modulation of Stem Cells as Therapeutics for Severe Mental Disorders and Cognitive Impairments. Front Psychiatry. 2020;11:80. doi:10.3389/fpsyt.2020.00080
- Ye N, Li B, Mao Q, et al. Orphan Receptor GPR88 as an Emerging Neurotherapeutic Target. ACS Chemical Neuroscience. 2019/01/16 2019;10(1):190-200. doi:10.1021/acschemneuro.8b00572
- Chen G, Xu J, Inoue A, et al. Activation and allosteric regulation of the orphan GPR88-Gi1 signaling complex. Nature communications. 2022/05/02 2022;13(1):2375. doi:10.1038/s41467-022-30081-5
- Del Zompo M, Deleuze JF, Chillotti C, et al. Association study in three different populations between the GPR88 gene and major psychoses. Mol Genet Genomic Med. Mar 2014;2(2):152-9. doi:10.1002/mgg3.54
- Sargazi S, Heidari Nia M, Saravani R, Jafari Shahroudi M, Jahantigh D, Shakiba M. IGF2BP2 polymorphisms as genetic biomarkers for either schizophrenia or type 2 diabetes mellitus: A case-control study. Gene Reports. 2020/09/01/ 2020;20:100680. doi:10.1016/j.genrep.2020.100680
- Sánchez-Romo D, Hernández-Vásquez CI, Pereyra-Alférez B, García-García JH. Identification of potential target genes in Homo sapiens, by miRNA of Triticum aestivum: A cross kingdom computational approach. Noncoding RNA Res. Jun 2022;7(2):89-97. doi:10.1016/j.ncrna.2022.03.002
- Bousman CA, Chana G, Glatt SJ, et al. Preliminary evidence of ubiquitin proteasome system dysregulation in schizophrenia and bipolar disorder: Convergent pathway analysis findings from two independent samples. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics. 2010/03/01 2010;153B(2):494-502. doi:10.1002/ajmg.b.31006
- Andrews JL, Goodfellow FJ, Matosin N, et al. Alterations of ubiquitin related proteins in the pathology and development of schizophrenia: Evidence from human and animal studies. Journal of Psychiatric Research. 2017/07/01/ 2017;90:31-39. doi:10.1016/j.jpsychires.2017.01.009
- Bousman CA, Luza S, Mancuso SG, et al. Elevated ubiquitinated proteins in brain and blood of individuals with schizophrenia. Scientific reports. Feb 19 2019;9(1):2307. doi:10.1038/s41598-019-38490-1
- Luza S, Opazo CM, Bousman CA, Pantelis C, Bush AI, Everall IP. The ubiquitin proteasome system and schizophrenia. The Lancet Psychiatry. 2020/06/01/ 2020;7(6):528-537. doi:10.1016/S2215-0366(19)30520-6
- McKinney BA, Reif DM, Ritchie MD, Moore JH. Machine Learning for Detecting Gene-Gene Interactions. Applied Bioinformatics. 2006/06/01 2006;5(2):77-88. doi:10.2165/00822942-200605020-00002
- Purcell SM, Wray NR, Stone JL, et al. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature. Aug 6 2009;460(7256):748-52. doi:10.1038/nature08185
- MacLean CJ, Sham PC, Kendler KS. Joint linkage of multiple loci for a complex disorder. Am J Hum Genet. Aug 1993;53(2):353-66.
- Hoh J, Ott J. Scan statistics to scan markers for susceptibility genes. Proc Natl Acad Sci USA. 2000;97(17):9615-7. doi:10.1073/pnas.170179197
- Hoh J, Ott J. Scan statistics in genome-wide scan for complex trait loci. In: Glaz J, Pozdnyakov V, Wallenstein S, eds. Scan Statistics: Methods and Applications (Statistics for Industry and Technology). Birkhäuser Boston; 2009:197-204.
- Hoh J, Wille A, Ott J. Trimming, weighting, and grouping SNPs in human case-control association studies. Genome Res. 2001;11(12):2115-9. doi:10.1101/gr.204001
- Landi I, Kaji DA, Cotter L, et al. Prognostic value of polygenic risk scores for adults with psychosis. Nature Medicine. 2021/09/01 2021;27(9):1576-1581. doi:10.1038/s41591-021-01475-7
- Barbu MC, Viejo-Romero M, Thng G, et al. Pathway-Based Polygenic Risk Scores for Schizophrenia and Associations With Reported Psychotic-like Experiences and Neuroimaging Phenotypes in the UK Biobank. Biological Psychiatry Global Open Science. 2023/10/01/ 2023;3(4):814-823. doi:10.1016/j.bpsgos.2023.03.004