





Effects of Spirulina on Antioxidant Systems in Poultry
Effects of a low protein soybean meal diet with and without Spirulina platensis freshwater microalgae on antioxidant systems in broiler liver and muscle tissue
Kentu Lassiter¹*, Garrett Mullenix², Sam Rochell³, Guillermo Tellez⁴, Michale T. Kidd¹, Sami Dridi¹, and Walter Bottje¹
- Center of Excellence for Poultry Science, Division of Agriculture, University of Arkansas, Fayetteville 72701, USA.
- Cargill Corporation, Minneapolis, MN55440, USA.
- Auburn University, Dept. of Poultry Science, Auburn, AL 36849, USA.
- Consultant
OPEN ACCESS
PUBLISHED: 30 November 2024
CITATION: Lassiter, K., Mullenix, G., et al., 2024. Effects of a low protein soybean meal diet with and without Spirulina platensis freshwater microalgae on antioxidant systems in broiler liver and muscle tissue. Medical Research Archives, [online] 12(11).https://doi.org/10.18103/mra.v12i11.5697
COPYRIGHT: © 2024 European Society of Medicine. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
DOI https://doi.org/10.18103/mra.v12i11.5697
ISSN 2375–1924
ABSTRACT
As the world’s population continues to increase and resources become more limited, sustainability and improved efficiency in poultry production has taken center stage across the industry. One specific issue facing the poultry industry is providing a cost-effective source of protein in broiler diets. Currently, soybean meal is the preferred choice of protein in broiler diets due to its relatively low cost. However, global human population growth, and the resulting increase in the demand for animal protein has begun to impact the price and availability of soybean meal. Furthermore, using soybean for animal feed comes with its own issues as protein sources for livestock, including the presence of anti-nutritional factors, and the plant protein digestibility. This study aims to investigate the effects of a low protein soybean meal diet supplemented with Spirulina platensis on antioxidant systems in broiler liver and muscle tissue.
Keywords
Spirulina platensis, soybean meal, broiler, antioxidant systems, muscle tissue
Introduction
As the world’s population continues to increase and resources become more limited, sustainability and improved efficiency in poultry production has taken on greater interest across the industry. One specific issue facing the poultry industry is providing a cost-effective source of protein in broiler diets. Currently, soybean meal is the preferred choice of protein in broiler diets due to its relatively low cost. However, global human population growth, and the resulting increase in the demand for animal protein has begun to impact the price and availability of soybean meal. Furthermore, using soybean for animal feed competes with its use as a protein source for humans. This issue has led to the search for alternative protein sources (which maintain or improve growth performance and overall bird health) that can be used in poultry diets.
A number of reports have indicated that Spirulina (Arthrospira) platensis is a promising candidate for use as a protein source in animal feed. Spirulina is a blue-green microalgae (cyanobacteria) that naturally grows in marine or freshwater aquatic systems and has good nutritional value due to its high protein content, and a good balance of essential amino acids and fatty acids¹ ². Earlier studies investigating the use of Spirulina as a feed supplement in livestock animals (e.g.³, including chickens, pigs, cattle, sheep, and rabbits) indicate that the algae potentially offers an improvement in health, production, and meat quality of the animals. A positive attribute of Spirulina in terms of contributing to sustainable poultry feed production is that it can be produced under a variety of environmental conditions with minimal land requirements; whereas soybeans are best suited for growth in (sub-) tropical climates, require more land for cultivation, and have a larger environmental footprint⁴. In addition to its nutritional value and sustainable production, Spirulina contains beneficial bioactive compounds such as phenolic acids, vitamins, minerals, and gamma linoleic acids⁵–⁷. Spirulina is also rich in pigments such as carotenoids, specifically xanthophylls and carotenes, and the plant protein phycocyanin, all of which contribute to excellent antioxidant/anti-inflammatory capabilities⁸ ⁹. These antioxidant/anti-inflammatory compounds contained in the algae are of particular interest as it relates to poultry production. The inclusion of Spirulina in poultry diets has been reported in a number of studies. Birds raised under conventional conditions have shown improved growth performance¹⁰ as well as improved antioxidant activity (Park et al.¹¹) when Spirulina is added to the diet. However, Bonos et al¹² did not show any effect of Spirulina on broiler performance. Recently, Mullenix et al.¹ also showed that the inclusion of Spirulina in low-protein broiler diets reduced the levels of pro-inflammatory cytokines in the circulation¹³. The benefits of algae inclusion on broiler performance and antioxidant status was apparent under heat stress conditions, where the induction of oxidative stress has a marked impact⁷. Heat stress increases the production of reactive oxygen species and impairs the mitochondrial species in the mitochondria of broiler skeletal muscle¹⁴ ¹⁵. Other studies reported that adding Spirulina to the diet of heat-stressed broilers improved and/or restored growth performance concomitant with increased antioxidant activity² ¹⁶ ¹⁷.
Numerous antioxidants work in concert in the cell to combat the constant production of oxidants coming from extracellular and intracellular sources (see review by Yu¹⁸). An overview of several redox coupled reactions in the cell is provided in Figure 1. This antioxidant/free radical scavenging milieu are often found in specific cellular compartments (e.g. cytosol and mitochondria) and can interact to rejuvenate each other by donating reducing equivalents. The major nonenzymatic antioxidant in the cell is reduced glutathione (GSH) which is found in mM concentrations in most cells. The rate limiting amino acid and enzyme for synthesis of GSH is cysteine and glutamate cysteine ligase (GCL), respectively. An interorgan circulation of GSH in which GSH is exported from the liver and taken up by other tissues, was first reported by Anderson et al.¹⁹ and later confirmed in broilers by Wang et al.²⁰. Thioredoxin is also an important thiol-containing antioxidant in the cell that helps convert numerous oxidized forms of antioxidants back to their reduced state (see review by Arner and Holmgren²¹) via redox reactions. The GSH recycling enzyme system consists of GSH peroxidase that reduces hydrogen and lipid peroxides using reducing equivalents from GSH to form oxidized glutathione (GSSG) that is recycled back to GSH by the action of GSH reductase using reducing equivalents from NADPH. Superoxide dismutase (SOD) catalyzes the reduction of superoxide to hydrogen peroxide. Isoforms of GSH peroxidase and SOD can be found within the cytosol and mitochondrial compartments.
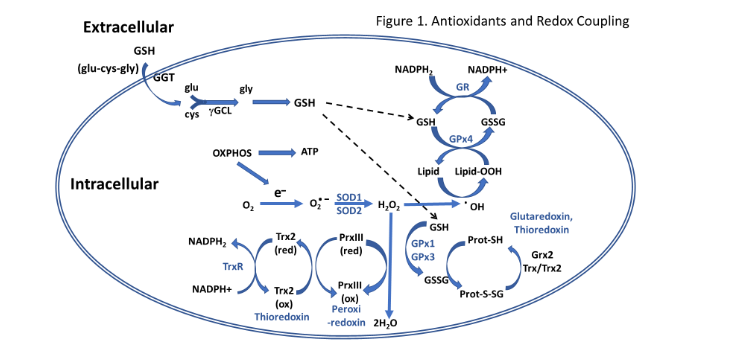
Figure 1: An overview of antioxidant systems and reduction-oxidation in the cell involved in free radical scavenging and antioxidant activity.
It is well documented that an increase in oxidative stress negatively impacts growth performance of broilers. In several studies comparing birds from the same genetic lineage, broilers with a low feed efficiency phenotype were shown to have higher levels of oxygen radicals and/or protein carbonyls (indicative of oxidative damage to proteins) in several tissues, including breast muscle²², heart muscle²³, liver²⁴ ²⁵, duodenum²⁶, and lymphocytes²⁷.
The inherently elevated metabolic activity in the liver and breast muscle of broilers means that these tissues are potentially more susceptible to oxygen radical production, and thus an increase in oxidative damage. The studies by Bonos et al.¹² and Park et al.¹¹ determined effects of Spirulina on blood antioxidant levels, but did not report effects on tissue antioxidants. It would be beneficial to also determine what antioxidant changes are occurring within the tissues as well to gain a more complete understanding of how including algae may be beneficial to broiler production. It should be noted that the breast muscle and liver tissues used in the current study were obtained from the same trial conducted and reported by Mullenix et al.¹. The purpose of this study was to determine if Spirulina supplementation in a low-protein broiler diet affected antioxidant status in breast muscle and liver tissues of male broilers, specifically looking at changes in antioxidant gene expression, as well as activities of key antioxidant enzymes, and nonenzymatic concentrations.
Materials and Methods
ANIMAL CARE STATEMENT
The humane care and treatment of animals were approved by the University of Arkansas Institutional Animal Care and Use Committee (IACUC) (IACUC protocol #21002).
ANIMALS AND TISSUES
Ross 708 male broilers were reared in floor pens and provided access to feed and water ad libitum. From day 1 to day 14, all birds were provided a standard commercial starter diet. From d 15 to 37, the birds were divided into three groups and provided one of three grower diets: 1) A grower diet (Positive control) consisting of a standard corn/soybean meal diet (CON), 2) a negative control, consisting of a low crude protein diet (LCP), and 3) the low crude protein diet in which Spirulina platensis meal was included at 10%, which represented half of the whole protein sources in the LCP plus microalgae (LCP+AL) diet as described by Muellenix et al.¹. Growth performance traits (weight gain, feed intake, feed conversion ratio, mortality) were reported in Muellenix et al. (2021). At the end of the trial, 10 birds were randomly selected from each treatment group, humanely euthanized by cervical dislocation, and breast muscle and liver tissues collected. The collected tissues were immediately frozen in liquid nitrogen and stored at –80°C until further analysis. Body weights were 2.61 ± 0.07, 2.60 ± 0.07, and 2.72 ± 0.08 for birds sampled from the CON, LCP, and LCP+AL, respectively.
RNA ISOLATION AND QUANTITATIVE REAL-TIME PCR
Total RNA was extracted from the breast muscle and liver tissues by TRIzol reagent (Life Technologies, Thermo Fisher Scientific, Carlsbad, CA) according to the manufacturer’s recommendations, DNase-treated, and reverse-transcribed (Quanta Biosciences, Gaithersburg, MD). The concentration and purity of RNA were determined for each sample using a Take 3 microvolume plate and a Synergy HT multimode microplate reader (BioTek, Winooski, VT). The reverse-transcription (RT) products (cDNAs) were amplified by real-time quantitative PCR (7500 realtime PCR system, Applied Biosystems, Thermo Fisher Scientific, Foster City, CA) with Power SYBR Green Master Mix (Applied Biosystems). Oligonucleotide primers used for chicken glutamate cysteine ligase catalytic subunit (GCLC), gamma-glutamyl transferase 2 (GGT2), glutathione peroxidases (GPx1, GPx3, and GPx4), glutathione reductase (GRd), thioredoxin reductase 1 (TrxR), Cu/Zn superoxide (SOD1) and MnSOD(SOD2), and the housekeeping gene ribosomal 18S are summarized in Table 1. The quantitative PCR (qPCR) cycling conditions were 50°C for 2 min, 95°C for 10 min followed by 40 cycles of a two-step amplification program (95°C for 15 s and 58°C for 1 min). At the end of the amplification, melting curve analysis was applied using the dissociation protocol from the Sequence Detection system to exclude contamination with unspecific PCR products. Relative expressions of target genes were determined by the 2^–ΔΔCt method²⁸.
Table 1
| Gene | Accession Numberᵃ | Primer Sequence (5’ → 3’) | Orientation | Product Size, bp |
|---|---|---|---|---|
| GCLC | XM_040666478.1 | TATGGGGGAACCACTGTCGGA (Forward) / CGGGACACCCTAACCTTGTGA (Reverse) | Forward / Reverse | 133 |
| GGT2 | XM_046901324.1 | GCTCTCGGAAAGAGGAGACTT (Forward) / CCATGAGCTITTGTGTGCAG (Reverse) | Forward / Reverse | 76 |
| GPx1 | NM_001277853.2 | TCCCGCTGAAACCAATTCG (Forward) / AGGGCAGGAGTCTCCTCGTT (Reverse) | Forward / Reverse | 57 |
| GPx3 | NM_001136232.2 | GGGCGCTGACCATCGAT (Forward) / CATCTTCCCCGGGGAAT TTC (Reverse) | Forward / Reverse | 59 |
| GPx4 | NM_001346448.1 | AGAATGGCGGAGCAGAGTG (Forward) / ATGGACAAAGAGGCCCTCTA (Reverse) | Forward / Reverse | 107 |
| GR | XM_040671422.1 | AGGAAGCAGCAGAGGTCAGG (Forward) / CTTCACATCGTGTGTCCTGG (Reverse) | 97 |
| Gene | Accession Numberᵃ | Primer Sequence (5’ → 3’) | Orientation | Product Size, bp |
|---|---|---|---|---|
| SOD1 | NM_205064.1 | TGGCTTCCATGTGCATGAAT (Forward) / AGCACCTGGCCTGGTACAC (Reverse) | Forward / Reverse | 58 |
| SOD2 | NM_204211.1 | GCTGGAGCCCCACATCAGT (Forward) / GGTGGGCTGGTGTTTGGCT (Reverse) | Forward / Reverse | 61 |
| TrxNRD1 | NM_001030762.2 | AGAGCACTGACCCAGCTTTATT (Forward) / GTGTGAAGAGGAAGCCCTGAATC (Reverse) | Forward / Reverse | 126 |
| 18S | AF173612 | TCCCCTCCCGTTACTTGAT (Forward) / GCGCTCGTCTGGCATGTA (Reverse) | Forward / Reverse | 60 |
ᵃ Accession numbers refer to GenBank (National Center for Biotechnology Information).
GLUTATHIONE ANALYSIS
Analysis of glutathione in breast muscle and liver tissues was conducted using a glutathione assay kit (#703002, Cayman Chemical, Ann Arbor, MI) following the manufacturer’s recommended procedure. The assay was used to measure total glutathione (TGSH), which is the sum of reduced glutathione (GSH) and oxidized glutathione (GSSG)²⁹. The assay is based on the enzymatic recycling method originally described by Tietze³⁰. The expression of TGSH and GSSG is reported as μM/mg protein.
GLUTATHIONE PEROXIDASE ANALYSIS
Analysis of glutathione peroxidase (GPx) activity in breast muscle and liver tissues was conducted using a GPx assay kit (#703102, Cayman Chemical, Ann Arbor, MI) following the manufacturer’s recommended procedure. The assay is based on the spectrophotometric assay described by Paglia and Valentine³¹. The activity of GPx is reported as nmol/min/mg protein.
GLUTATHIONE REDUCTASE ANALYSIS
The measurement of glutathione reductase (GRd) activity in breast muscle and liver tissues was performed using a GRd assay kit (#703202, Cayman Chemical, Ann Arbor, MI) following the manufacturer’s recommended procedure. GRd activity is reported as nmol/min/mg protein.
SUPEROXIDE DISMUTASE ANALYSIS
Analysis of superoxide dismutase (SOD) activity in breast muscle and liver tissues was conducted using a SOD assay kit (#706002, Cayman Chemical, Ann Arbor, MI) following the manufacturer’s recommended procedure. The expression of SOD activity is reported as U/mg protein.
THIOREDOXIN REDUCTASE ANALYSIS
The measurement of thioredoxin reductase (TrxR) activity in breast muscle and liver tissues was performed using a TrxR assay kit (#10007892, Cayman Chemical, Ann Arbor, MI) following the manufacturer’s recommended procedure. TR activity is reported as nmol/min/mg protein.
THIOBARBITURIC ACID REACTIVE SUBSTANCES (TBARS) ANALYSIS
Measurement of the lipid peroxidation product malondialdehyde (MDA) in breast muscle and liver tissues was conducted using a TBARS assay kit (#10009055, Cayman Chemical, Ann Arbor, MI) following the manufacturer’s recommended procedure. The expression of MDA is reported as nM/mg tissue.
STATISTICAL ANALYSES
In the qPCR and oxidative assay studies, comparison of mean expression values between the three experimental diets were made using one way analysis of variance (ANOVA) and student t-test with Graph Pad Prism (version 7.0). Differences were considered significant at P ≤ 0.05.
Results
LIVER
In the liver, mRNA expression of γGT, the enzyme needed for hydrolysis of GSH and transport into the cells, was highest in the LCP+AL group and lowest in the CON group (Figure 2A). The expression of GCL, the rate limiting enzyme in GSH synthesis, was elevated (P<0.05) in the LCP-AL group compared to CON-fed and LCP fed birds (Figure 2B). Levels of hepatic TGSH were highest in the CON-fed birds and lowest in birds receiving the LCP diet (Figure 2C). Both GSSG and TBARS, (indices of oxidative stress in the cytosolic and lipid compartments, respectively), were higher in the LCP-AL group compared to the LCP group (Figure 2D and 2E). The percentage of GSSG in TGSH (insert, Figure 2D) was 0.06% for CON, 0.04% for LCP groups, and 0.2% for the LCP AL group. There was no difference in mRNA expression in SOD1 (CuZn SOD) between treatment groups (Figure 2F). Whereas SOD2 expression was higher in the LCP+AL group compared to CON values (Figure 2G), overall SOD activity was highest in CON and lowest in the LCP AL group (Figure 2H). The mRNA expression of TrxR was higher in the LCP+AL group compared to CON (Figure 2I), but there were no differences in TrxR enzyme activity (Figure 2J). Expression and activity of GSH recycling enzymes are shown in Figure 2K to 2P. In general, the highest expression was observed in the LCP AL group, and lowest values observed in the CON group. Liver activity of GPx was lower in the LCP AL group compared to both the CON-fed and LCP-fed (GPx) (Figure 2N). There was a marginal reduction of Grd activity (P = 0.09) in the LCP+AL group compared to the LCP group (Figure 2P)
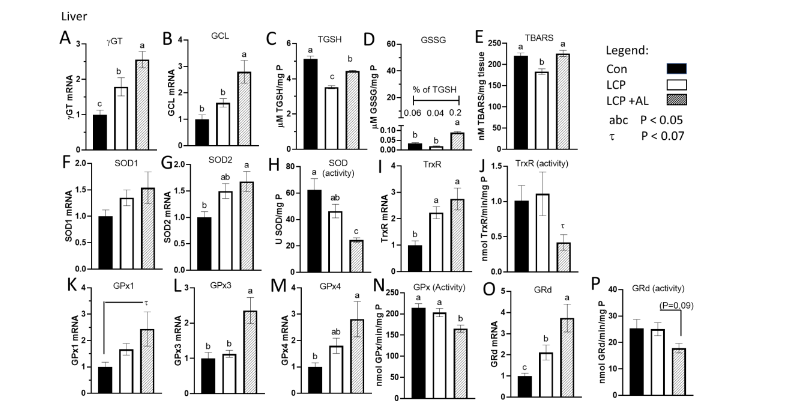
Figure 2. Effect of feeding Spirulina platensis microalgae on antioxidant gene expression and enzyme activities in liver. Broilers were either a standard corn-soy based Control (CON) diet (20% CP), a low crude protein diet (LCP, 17% CP), or the LCP diet with half of the soybean meal (10% of the diet) was replaced with microalgae (LCP + AL, 17% CP). Relative gene expression is presented in arbitrary units compared to values obtained in the CON diet, concentrations of total GSH (TGSH) and oxidized GSH (GSSG) are presented in μmole per mg P, and enzyme activities are presented in units per mg tissue P. Data are presented for: A) gamma-glutamyl transpeptidase (γGT), B) glutamate cysteine ligase (GCL), C) Total GSH (TGSH), D) oxidized glutathione (GSSG), E) thiobarbituric acid reducing substance (TBARS), F) superoxide dismutase 1 (SOD1), G) SOD2, H) SOD enzyme activity, I) thioredoxin (TrxR), J) TrxR enzyme activity, K) GSH peroxidase 1 (GPx1), L) GPx3, M) GPx4, N) GPx enzyme activity, O) glutathione reductase (GRd), and P) GRd enzyme activity.
Data are presented as mean ± SEM (n = 7–10/group).
ᵃᵇᶜ Means with different letters are significantly different (P < 0.05).
BREAST MUSCLE
The expression of γGT was higher in the LCP+AL-fed birds compared to the CON and LCP groups (Figure 3A) whereas as GCL expression was lower (P = 0.067) in the LCP and LCP+AL groups compared to CON (Figure 3B). Total GSH and GSSG levels were higher in breast muscle of the LCP AL compared to the CON and LCP groups (Figure 3C and 3D). There were no differences in TBARS between groups (Figure 3E). Whereas expression of SOD1 (CuZn) was marginally lower (P< 0.09) in the LCP and LCP AL birds (Figure 3F), SOD2 (Mn) was lower in the LCP+AL compared to the other two groups (Figure 3G). Overall SOD activity was higher in muscle of the LCP+AL-fed birds compared to CON (Figure 3H). The LCP+AL-fed birds exhibited higher TrxR expression and activity compared to the other two groups.
Analysis of glutathione in breast muscle and liver tissues was conducted using a glutathione assay kit (#703002, Cayman Chemical, Ann Arbor, MI) following the manufacturer’s recommended procedure. The assay was used to measure total glutathione (GSH) and oxidized glutathione (GSSG). groups (Figure 3I and 3J). With respect to GPx, only GPx3 was elevated in the LCP+AL group, but there were no treatment differences in GPx1 and GPx4 expression or GPx activity (Figure 3K–N). Both GRd expression (Figure 3O) and activity (Figure 3P) were higher in the LCP+AL group compared to the CON but were not different to the LCP values.
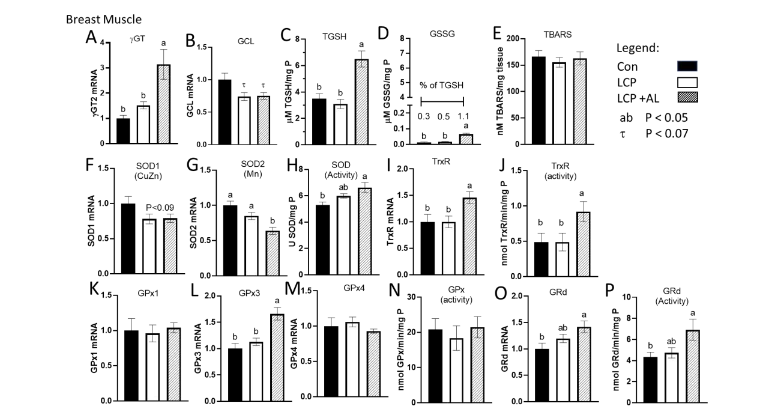
Figure 3. Effect of feeding Spirulina platensis microalgae on antioxidant gene expression and enzyme activities in breast muscle. Broilers were either a standard corn-soy based Control (CON) diet (20% CP), a low crude protein diet (LCP, 17% CP), or the LCP diet in which half of the soybean meal (10% of the diet) was replaced with microalgae (LCP + AL, 17% CP). Relative gene expression is presented in arbitrary units compared to values obtained in the CON diet, concentrations of total GSH (TGSH) and oxidized GSH (GSSG) are presented in μmole per mg P, and enzyme activities are presented in units per mg tissue P. Data are presented for: A) gamma-glutamyl transpeptidase (γGT), B) glutamate cysteine ligase (GCL), C) Total GSH (TGSH), D) oxidized glutathione (GSSG), E) thiobarbituric acid reducing substance (TBARS), F) superoxide dismutase 1 (SOD1), G) SOD2, H) SOD enzyme activity, I) thioredoxin (TrxR), J) TrxR enzyme activity, K) GSH peroxidase 1 (GPx1); L) GPx3, M) GPx4, N) GPx enzyme activity, O) glutathione reductase (GRd), and P) GRd enzyme activity.
Data are presented as mean ± SEM (n = 7–10/group).
ᵃᵇᶜ Means with different letters are significantly different (P < 0.05).
Discussion
We previously reported that feeding a low crude protein (LCP) diet in which half of the soybean meal was replaced by microalgae (Spirulina platensis) meal (LCP+AL), had no adverse effect on broiler growth performance (comparing the LCP and LCP+AL diets)¹. Crafton³² (2022) also reported no difference in growth performance in broilers fed diets containing 2% microalgae with or without 8% distiller dried grains. Birds fed the LCP+AL diet exhibited reductions in bacterial translocation from the gut, food pad lesions, as well as circulating cytokines, and an increase in pigmentation of skin and muscle¹. The increase in pigmentation was most likely due to higher levels of carotenoids in the LCP+AL diet and could be hypothesized to reduce oxidative stress. Spirulina platensis microalgae is a very good source of antioxidants and thus can be expected to be beneficial in helping combat cellular oxidative stress (e.g. Kumar et al.³³). Oxidative stress occurs when the levels of free radical production overcome the host’s physiological ability to effectively scavenge them, ultimately resulting in DNA, protein, and lipid damage. Thus, a major goal of the current study was to investigate the impact of a low crude protein (LCP) diet (17% vs 21% crude protein in a control diet, [CON]), and one in which half of the LCP soybean meal was replaced with Spirulina platensis (LCP+AL) on antioxidant systems (gene expression, enzyme activity and tissues (liver and breast muscle) obtained in male broilers.
attenuates lipid peroxidation and DNA damage¹³ ¹⁴ ³⁹. Analysis of Spirulina indicates that the pigment protein phycocyanin and the vitamin A precursor β-carotene are the two primary active compounds in the microalgae, with both taking part in antioxidant and anti-inflammatory activities (reviewed by Wu et al.⁴⁰). Reports have shown that phycocyanin effectively decreases lipid peroxidation, the formation of peroxyl, alkoxyl, and hydroxyl radicals, nitrite production, and iNOS expression⁴¹ ⁴². β-carotene has also been characterized as having antioxidant effects by inhibiting the intracellular accumulation of reactive oxygen species and protecting membranes against singlet oxygen-mediated lipid peroxidation¹⁸ ⁴³. Anti-inflammatory activity of β-carotene in mice macrophages stimulated by lipopolysaccharide (LPS) or interferon gamma (IFNγ) is due to down-regulation of several genes associated with inflammation (COX-2, TNF-α, IL-1β, IL-6, and IL-12) that is likely due to inhibition of NF-KB and iNOS promoters⁴⁴ ⁴⁵.
There was a decrease in TGSH and GSH (not shown) in the liver in the LCP-fed birds compared to CON-fed birds, that was partially ameliorated in birds fed the LCP+AL diet (Figure 2C). However, there was also evidence of increased oxidative stress as indicated by the increase in GSSG and TBARS (Figure 2D and 2E). There were also elevations in TGSH and GSSG in breast muscle of LCP+AL birds, but no difference in TBARS (representing generalized lipid oxidation) compared to LCP-fed (Figure 3C, D, and E). It should be noted that despite the elevation in GSSG in liver and muscle, the ratio of TGSH and GSSG was 99.8 in liver and 98.9 in breast muscle and on the low end of an acceptable range of 1 to 5%¹⁸.
This should not be unexpected since the liver is continuously exposed to oxidative stress conditions arising from: 1) the detoxification of xenobiotics and other substances that have been absorbed in the intestines and delivered to the liver via the portal vein, and 2) the substantial and continuous production of ROS from hepatic mitochondria that must meet the high energy requirements of the organ. Analysis of multiple components of the glutathione system was conducted to determine how gene expression and enzyme activity was altered in response to microalgae. In addition to its vital role as a participant in antioxidant defense and redox homeostasis, the glutathione system is also a key part of cell signaling, cellular metabolism, and immunomodulation. The peptide glutathione (GSH) is an essential antioxidant in that it detoxifies reactive oxygen species (ROS), reactive nitrogen species (RNS), and hydroxyl radicals. The liver is one of the organs with the highest GSH content but interestingly, gene expression for glutamate cysteine ligase (GCL), the rate-limiting enzyme in GSH synthesis, was significantly increased in the liver of birds fed the LCP+AL diet; however, GCL gene expression was significantly lower in the breast muscle of LCP+AL birds. This difference in GCL expression between the two tissue types may be partially explained by the interorgan transfer of GSH²⁰. It is possible that supplementation of the broiler diet with Spirulina is further stimulating gene expression of enzymes involved in glutathione synthesis in the liver (i.e. increasing GCL, GPx, GRd gene expression), which is followed by the increased production and exportation of GSH into the general circulation and uptake by tissues (e.g. muscle) for resynthesis. Previous work by Wang et al.²⁰ has demonstrated the ability of the avian liver to export GSH into the general circulation. Since GCL activity is regulated by GSH via a negative feedback loop⁴⁶, an increase in GSH activity (as observed in this study) in the breast muscle through interorgan transfer from the liver may be at least partially responsible for the reduced GCL expression observed in the breast muscle of broilers fed the LCP+AL diet. GSH also serves as a cofactor for enzymes in the GSH recycling system⁴⁷ ⁴⁸. There are multiple isozymes of GPx that have been identified, whose function is to catalyze the reduction of hydroperoxides to corresponding alcohols, and H₂O₂ to water using reduced glutathione as an electron donor⁴⁹.
relation to the present study, GPx1 is the most abundant isozyme and found in the cytosol, mitochondria, and nucleus of most tissues; whereas GPx3 is a secreted form found in plasma in various organs, and GPx4 (i.e. phospholipid GPx) is a membrane-bound isozyme found in the cytosol, mitochondria, and nucleus of different tissues⁵⁰ ⁵¹. Gene expression levels of GPx1, GPx3, and GPx4 were observed in breast muscle and liver tissues collected during the present study. In liver tissue, expression of all three GPx isozymes was up-regulated in birds fed the LCP+AL diet when compared to the Standard diet, however only GPx3 was up-regulated in breast muscle. Expression of the gene encoding glutathione reductase (GRd), which is the enzyme responsible for catalyzing the reduction of oxidized glutathione (GSSG) to GSH was significantly increased in both tissue types of birds fed the LCP+AL diet. GRd is very important to preserving a healthy redox balance since its activity is responsible for replenishing and maintaining a high ratio of GSH to GSSG⁹ ²⁷. Similar to GRd, gene expression of the enzyme thioredoxin reductase (TrxR), whose actions regenerate reduced thioredoxin in the antioxidant system, was up-regulated in both breast muscle and liver tissue. Both GRd and TrxR are important components in the regulation of redox status and maintenance of redox homeostasis. The suggestion that the glutathione and thioredoxin systems work in concert during the scavenging of H₂O₂ ⁵³ would help explain the similar patterns seen in gene expression of the two reductases following dietary treatment with microalgae in the current study.
In addition to investigating the GSH system, superoxide dismutase (SOD) gene expression following the inclusion of microalgae in the diet was also studied. Superoxide is the most-prevalent free radical under normal cellular physiological conditions, therefore SOD is an important antioxidant and is the primary contributor to the first line of antioxidant defense against ROS formation and subsequent damage to DNA, lipids, proteins, and carbohydrates⁵⁴ ⁵⁵. In the present study there was no difference in relative mRNA expression of SOD1 in liver and breast muscle tissue between the three diets; however, expression of SOD2 in the LCP+AL-fed birds was significantly up-regulated in liver, but significantly down-regulated in the breast muscle, further supporting the observations that there are tissue-specific differences in the avian antioxidant defense system. In fact, inherent tissue-specific differences in the chicken antioxidant profile have been previously reported⁵⁶. Further investigation of the specific microalgae Spirulina platensis reveals some possible explanations for the obtained results. As stated earlier, Spirulina platensis contains carotenoids, a class of natural pigments that serve as a major dietary source of vitamin A, participates in the scavenging of free radicals, and are further divided into two groups consisting of xanthophylls and carotenes based on their chemical structure⁵⁶. When these compounds are incorporated into the diet, they are absorbed into the body through passive diffusion via intestinal epithelial cells and metabolized⁵⁷. In addition to its scavenging properties, reports have indicated that carotenoids activate the network of antioxidant defenses by interacting with transcription factors that regulate it. The redox-sensitive transcription factor Nrf2 (nuclear factor-erythroid factor 2-related factor 2) is of particular interest since it is considered the master regulator in adaptive oxidative stress responses⁵⁸. The activation of Nrf2 results in it translocating to the nucleus where it binds to the antioxidant response element (ARE) and induces the transcription of several antioxidant enzymes, including GCL, GPx, GRd, SOD, TrxR⁵⁹. Interestingly, dietary regulation of the antioxidant pathway involving Nrf2 and ARE has been discussed previously. The review by Stefanson and Bakovic⁶⁰ cites a number of dietary phytochemicals that have been shown to upregulate the transcription of antioxidant enzymes in the absence of oxidative stress. These include many plant-based dietary phytochemicals which would include carotenoids ⁶¹ ⁶². Therefore, an increase in the presence of carotenoids in the diet and the
subsequent activation of the Nrf2 pathway would give a possible explanation as to why a number of antioxidant genes were upregulated (in the absence of oxidative stress induction) in the tissues of birds fed the LCP+AL diet.
In this study the inclusion of Spirulina platensis in a low crude protein broiler diet also affected antioxidant and antioxidant enzyme activity levels. Once again, the effects of the microalgae give rise to tissue-specific differences that will be discussed below. The results obtained in the liver tissue are peculiar, given that in the LCP+AL-fed birds the expression of several genes involved in glutathione synthesis and recycling were significantly upregulated, but there was no change in glutathione levels and the activity of nearly every enzyme measured was significantly reduced. The reduction in liver SOD, GPx, GR, and TrxR activity may well be attributed to the increased radical scavenging ability provided by Spirulina’s endogenous antioxidants (i.e., carotenoids, flavonoids, phycocyanins). It must also be considered that Spirulina platensis contains its own antioxidant defense system that is used to control ROS levels and protect cells from oxidative damage that occurs from environmental stress or the photosynthetic process. Analysis of the algae has shown that it produces several antioxidant enzymes including SOD¹⁷ ³³ ⁶³ as well as peroxidase, catalase, and ascorbate peroxidase⁶⁴. It appears that there is the potential for several different antioxidant enzymes to be absorbed and metabolized from the algae in a supplemented diet, ultimately leading to a reduction in the activity of host antioxidant enzymes as seen in the liver tissue. In contrast, in the breast muscle tissue a significant increase in antioxidant (total GSH and reduced GSH) and antioxidant enzyme activity (GRd, SOD, TrxR) was observed in birds fed LCP+AL. The differences in expression and activity between the two tissues cannot be fully explained at this time and further investigation is indeed warranted. However, one possible consideration for the differences between the two tissues reverts back to the previously discussed Nrf2 pathway. The modern broiler chicken has been genetically selected for high feed efficiency and rapid muscle growth in order to meet consumer demands. A global gene expression study conducted by Kong et al⁶⁵ comparing the breast muscle from a modern broiler line to an unselected foundation broiler line reported that the Nrf2 pathway is more active in modern broilers; the reasoning being that low ROS levels resulting from increased antioxidant defenses are necessary to sustain rapid muscle growth. Subsequent microRNA profiling of breast muscle from the same two broiler lines also indicated that Nrf2-mediated oxidative stress response pathways are activated for enhanced ROS scavenging in the rapidly growing muscles of modern broilers⁶⁶. This would suggest that the genetic selection for breast muscle growth present in modern broiler lines may produce an altered Nrf2 pathway response that would be different from what is observed in liver tissue.
One interesting result from this study that should be addressed is the increase in oxidized glutathione (GSSG) observed in both tissues of birds from the LCP+AL group, indicating that there may be increased oxygen radical production being addressed by the glutathione system. Although carotenoids such as β-carotene are known for their antioxidant activity, a number of studies have indicated that these compounds may also function as a prooxidant as well under certain conditions. The prooxidant potential of carotenoids⁶⁷ becomes more favorable in the face of increasing oxygen partial pressure (pO₂) and/or increasing carotenoid concentrations in tissues, resulting in more favorable conditions for autooxidation that leads to the formation of carotenoid alkoxyl/peroxyl radicals⁶⁸. Also, studies conducted in rats and mice indicate that the accumulation of carotenoids in tissues can disturb electron transport in mitochondria during ATP production, resulting in the generation of ROS⁶⁹ ⁷⁰. However, it is interesting to note that birds may be more resistant to the prooxidant effects of carotenoids than
mammals as suggested by Johnson and Hill⁷¹. Mayne and Parker⁷² reported that the accumulation of dietary beta-carotene in chicken liver is also accompanied by increased levels of the antioxidants α-tocopherol and retinol, potentially providing a pathway to arrest carotenoid pro-oxidation. Also, birds have a lower basal rate of electron leak and subsequent ROS formation compared to mammals⁷³. Therefore, it should be noted that although there was an increase in breast muscle and liver GSSG in birds fed the algae-supplemented diet, the ratio of GSH to GSSG (an indicator of oxidative stress) did not deviate outside of what would be considered a “normal” physiological range for a healthy redox balance (unpublished observations). In addition, there was no change in measured MDA levels (an indicator of lipid peroxidation) in breast muscle and liver tissue from the LCP+AL diet when compared to the Control diet. This antioxidant/prooxidant capability of carotenoids is interesting and may help to provide further insight into the tissue-specific results seen in this study. Experiments in both rats and birds where animals were fed carotenoid-supplemented diets reported that carotenoids accumulate and are stored in the liver⁶⁹ ⁷⁴ ⁷⁵. Different levels of carotenoid accumulation between the liver tissue and breast muscle tissue may provide further explanation for the observed tissue-specific differences in antioxidant responses; however, tissue carotenoid levels were not examined in this study and future work looking at the antioxidant properties of microalgae may warrant further investigation into how distribution of carotenoids among tissues affects antioxidant responses in those tissues.
Conclusion
The results from this study indicate that the supplementation of a low crude protein diet with Spirulina platensis protein meal enhances the antioxidant status of breast muscle and liver tissue in male broilers. Breast muscle showed an increase in GPx3, GRd, TrxR gene expression and an increase in GSH, GRd, SOD, and TrxR activity. In general, the expression of GPx1, GPx3, GPx4, GRd, SOD2, and TR was upregulated, while there was a significant reduction in the activity of several antioxidant enzymes (GPx, GRd, SOD, TrxR). It is proposed here that the inclusion of microalgae in the diet directly protects against oxidative stress by increased radical scavenging due to compounds originating from the microalgae (i.e. phycocyanins, carotenoids, flavonoids, and other vitamins). The direct scavenging of radicals via the microalgae may explain why there was a reduction in antioxidant enzyme activity in the liver tissue. Spirulina platensis may also enhance antioxidant protection in an indirect fashion through the activation of the antioxidant transcription factor Nrf2 and the subsequent antioxidant response element (ARE) to induce gene expression of antioxidant enzymes. However, the increase in GSSG seen in both tissues suggests that the inclusion of microalgae may become a concern due to the prooxidant potential of β-carotene. The antioxidant defense pathway of organisms is complex and interacts with a number of other pathways (i.e. immunomodulatory, anti-inflammatory) that may affect its actions. Also, other factors such as interorgan circulation of glutathione originating from the liver, the differences in metabolism of various tissues, and genetic selection for muscle growth must be taken into consideration as well. As a result, the differences in gene expression and enzyme activity observed between the two tissue types should not be unexpected and indeed requires further investigation. The inclusion of Spirulina platensis into poultry diets as an alternative protein source has benefits and should be pursued further.
Conflict of Interest:
The authors declare there are no competing interests.
Funding Statement:
None.
Acknowledgements:
This research was supported by funds provided by USDA NIFA Sustainable Agriculture Systems (#2019 69012-29905) and the Division of Agriculture, University of Arkansas.
Author Contributions:
Conceptualization; KL, GM, MK, SR, SD, WB
Methodology; KL, GM
Formal Analysis; KL, SD
Writing-original draft preparation; KL, WB, SD
All authors have read and agreed to this published version of the manuscript.
References
1. Mullenix, G. J. et al. Spirulina platensis Inclusion Reverses Circulating Pro-inflammatory (chemo) cytokine profiles in broilers fed low-protein diets. Front. Vet. Sci. 11, 1243 (2021).
2. Abdel-Moneim, A., Shehata, A. M., Mohamed, N. G., Elbaz, A. M. & Abrahim; NS. Synergistic effect of Spirulina platensis and selenium nanoparticles on growth performance, serum metabolites, immune responses, and antioxidant capacity of heat-stressed broiler chickens. Biol Trace Elem Res 200, 768–779 (2022).
3. Holman B.W. & Malau-Aduli A.E. Spirulina as a livestock supplement and animal feed. J. Anim. Physiol. Anim. Nutr. (Berl). . Anim. Physiol. Anim. Nutr. (Berl). 97, 615–623 (2013).
4. Taelman, S., De Meeste, S., Van Dijk, W., da Silva, V. & Dewulf, J. Environmental sustainability analysis of a protein-rich livestock feed ingredient in The Netherlands: Microalgae production versus soybean import. Resource Conserv Recycl 101, 61–72 (2015).
5. Becker EW. Microalgae: Biotechnology and Microbiology. (Cambridge University Press, 1994).
6. Kulshreshtha, A. et al. Spirulina in health care management. Curr. Pharm. Biotechnol. 9, 400–405 (2008).
7. Mariey, B., Samak, H. & Ibrahem, M. A. Effect of using Spirulina platensis algae as a feed additive for poultry diets: 1-Productive and reproductive performances of local laying hens. Egypt. Poult. Sci. J. 32, (2012).
8. Eriksen, N. T. Production of phycocyanin – a pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 80, 1–14 (2008).
9. Christaki, E., Bonos, E., Giannenas, I. & Florou-Paneri. Functional properties of carotenoids originating from algae. J. Sci. Food Agric. 93, 5–11 (2013).
10. Fathi, M. A., Namra, M. M., Ragab. M.S. & Aly, M. M. Effect of dietary supplementation of algae meal (Spirulina platensis) as growth promotor on performance on performance of broiler chickens. Egypt. Poult. Sci. 38:375–389. 38, 375–389 (2018).
11. Park, J. H., Lee, S. I. & Kim, I. H. Effect of dietary spirulina (arthrospira) platensis on the growth performance, antioxidant enzyme activity, nutrient digestibility, cecal microflora, excreta noxious gas emission, and breast meat quality of broiler chickens. Poult. Sci. 97, 2451–2459 (97AD).
12. Bonos, E. et al. Spirulina as a functional ingredient in broiler chicken diets. S. Afr. J. Anim. Sci. 46, 94–102 (2016).
13. Elbaz, A. M., Ahmed, A. M., Abdel-Maqsoud, A., Badran, A. M. & Abdel-Moneim, A. M. E. Potential ameliorative role of Spirulina platensis in powdered or extract forms against cyclic heat stress in broiler chickens. Environ. Sci. and Pollution Res. 29(30) 97, 45578–45588 (2022).
14. Mujahid A, Akiba Y & Toyomizu M. Olive oil-supplemented diet alleviates heat stress-induced mitochondrial ROS production in chicken skeletal muscle. Am. J. Phys. Regul. Integr. Comp. Physiol. 297, R690–R698 (2009).
15. Mujahid, A. et al. Mitochondrial oxidative damage in chicken skeletal muscle induced by acute heat stress. J. of Poultry Sci. 44, (2007).
16. Mirzaie S, Zirak-Khattab F, Hosseini SA & Donyaei-Darian H. Effects of dietary Spirulina on antioxidant status, lipid profile, immune response and performance characteristics of broiler chickens reared under high ambient temperature. Asian-Australas J. Anim. Sci. 31, 556–563 (2018).
17. Moustafa ES et al. Blue-green algae (Spirulina platensis) alleviates the negative Impact of heat stress on broiler production performance and redox status. Animals 11, 1243 (2021).
18. Yu, B. P. Cellular defenses against damage from reactive oxygen species. Physiol. Rev. 74, 139–162 (1994).
19. Anderson ME, Bridges RJ & Meister A. Direct evidence for interorgan transport of glutathione and that the non-filtration renal mechanism for glutathione utilization involves γ-glutamyl transpeptidase. Biochem. Biophys. Res. Comm 96, 848–853 (1980).
20. Wang, S. et al. Hepatic export of glutathione and uptake of constituent amino acids, glutamate and cysteine, in broilers in vivo. Poult Sci 77, (1998).
21. Amer, E. S. & Holmgren, A. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 81, 6102–6109 (2000).
22. Bottje, W. et al. Association of mitochondrial function with feed efficiency within a single genetic line of male broilers. Poult Sci 81, (2002).
23. Tinsley, N. et al. Investigation of mitochondrial protein expression and oxidation in heart muscle in low and high feed efficient male broilers in a single genetic line. Poult Sci 89, (2010).
24. Iqbal, M. et al. Low feed efficient broilers within a single genetic line exhibit higher oxidative stress and protein expression in breast muscle with lower mitochondrial complex activity. Poult Sci 83, (2004).
25. Iqbal, M. et al. Compromised liver mitochondrial function and complex activity in low feed efficient broilers are associated with higher oxidative stress and differential protein expression. Poult Sci 84, (2005).
26. Ojano-Dirain, C., Iqbal, M., Wing, T., Cooper, M. & Bottje, W. Glutathione and respiratory chain complex activity in duodenal mitochondria of broilers with low and high feed efficiency. Poult Sci 84, (2005).
27. Lassiter, K. et al. Differential expression of mitochondrial and extramitochondrial proteins in lymphocytes of male broilers with low and high feed efficiency. Poult Sci 85, (2006).
28. Schmittgen, T. D. & Livak, K. J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc 3, 1101–1108 (2008).
29. McGill, M. R. & Jaeschke, M. A direct comparison of methods used to measure oxidized glutathione in biological samples: 2-vinylpyridine and N-ethylmaleimide. Toxicol. Mech. Methods 25, 589–595 (2015).
30. Tietze, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: applications to mammalian blood and other tissues. Anal. Biochem. 27, 502–522 (1969).
31. Paglia, D. E. & Valentine, W. N. A direct comparison of methods used to measure oxidized glutathione in biological samples: 2-vinylpyridine and N-ethylmaleimide. J. Lab Clin. Med. 70, 158–169 (1967).
32. Crafton, S. W. Assessment of novel protein ingredient arthrospira platensis (microalgae) and soybean genotype amino acid and oil selection Improvements on broiler performance. (Division of Agriculture, University of Arkansas, 2022).
33. Kumar, A., et al. Antioxidants and phytonutrient activities of Spirulina platensis. Energy Nexus 6, 1–9 (2022).
34. Riss J et al. Phycobiliprotein C-phycocyanin from Spirulina platensis is powerfully responsible for reducing oxidative stress and NADPH oxidase expression induced by an atherogenic diet in hamsters. J. Agric. Food Chem. 55, 7962–7967 (2007).
35. Bermejo-Bescós P, Piñero-Estrada E & Villar del Fresno AM. Neuroprotection by Spirulina platensis protean extract and phycocyanin against iron-induced toxicity in SH-SY5Y neuroblastoma cells. Toxicol. In Vitro 22, 1496–1592 (2008).
36. Karadeniz A, Yildirim A, Simsek N, Kalkan Y & Celebi F. Spirulina platensis protects against gentamicin-induced nephrotoxicity in rats. Phytother. Res. 22, 1506–1510 (2015).
37. Koníčková R et al. Anti-cancer effects of blue-green alga Spirulina platensis, a natural source of bilirubin-like tetrapyrrolic compounds. Ann. Hepatol. 13, 273–283 (2014).
38. Abdelkhalek NK, Ghazy EW & Abdel-Daim MM. Pharmacodynamic interaction of Spirulina platensis and deltamethrin in freshwater fish Nile tilapia, Oreochromis niloticus: impact on lipid peroxidation and oxidative stress. Pollut. Res. Int. 22, 3023–3031 (2015).
39. Galip N, Seyidoglu N, Serdar Z & Savas N. . The effect of Saccharomyces Cerevisiae and Spirulina Platensis on glutathione and leucocytes count in rabbits. J. Res. Vet. Med 38, 71–76 (2019).
40. Wu Q et al. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: an overview. Arch. Toxicol. 90, 1817–1840 (2016).
41. Deng R & Chow TJ. Hypolipidemic, antioxidant, and antiinflammatory activities of microalgae Spirulina. Cardiovasc. Ther. 28, e33–e45 (2910).
42. Xia D et al. Protective effects of C-phycocyanin on alcohol-induced acute liver injury in mice. J. Oceanol. Limnol. 34, 399–404 (2016).
43. Schafer FQ et al. Comparing beta-carotene, vitamin E and nitric oxide as membrane antioxidants. Biol. Chem 383, 671–681 (2002).
44. Bai SK et al. Beta-Carotene inhibits inflammatory gene expression in lipopolysaccharide-stimulated macrophages by suppressing redox-based NF-kappaB activation. Exp. Mol. Med 37, 323–334 (2009).
45. Katsuura S, Imamura T, Bando N & Yamanishi, R. Beta-Carotene and beta-cryptoxanthin but not lutein evoke redox and immune changes in RAW264 murine macrophages. Mol. Nutr. Food. Res. 53, 1396–1405 (2009).
46. Lushchak, V. Glutathione homeostasis and functions: Potential targets for medical interventions. J. Amino Acids 2012/e736837., (2012).
47. Maher, P. The effects of stress and aging on glutathione metabolism. Aging Res. Rev. 288–314 (2005).
48. Vivancos Diaz P, Wolff T, Markovic J, Pallardo FV & Foyer CH. A nuclear glutathione cycle within the cell cycle. Biochem. J. 275, 169–178 (2010).
49. Margis R, Dunand C, Teixeira FK & Margis-Pinheiro M. Glutathione peroxidase family-an evolutionary overview. FEBS J. 275, 3959–3970 (2008).
50. Brown KM et al. Effects of organic and inorganic selenium supplementation on selenoenzyme activity in blood lymphocytes, granulocytes, platelets and erythrocytes. Clin. Scl. 98, 593–599 (2000).
51. Herbette S, Roeckel-Drevet P & Drevet JR. Seleno-independent glutathione peroxidases. FEBS J. 1830, 2163–2180 (2007).
52. Deponte, M. Glutathione catalysis and the reaction mechanisms of glutathione-dependent enzymes. Biochim. Biophys. Acta 1830, 3217–3266 (2013).
53. Aon MA et al. Glutathione/thioredoxin systems modulate mitochondrial H2O2 emission: an experimental-computational study. J. Gen. Physiol. 139, 479–491 (2012).
54. Halliwell, B. Free radicals and antioxidants: A personal view. Nutr. Rev 52, 253–265 (1994).
55. Surai PF, Kochish II, Fisinin VI & Kidd MT. Antioxidant defence systems and oxidative stress in poultry biology: An update. Antioxidants (Basel) 8, 235 (2019).
56. Jackson H, Braun CL & Ernst H. The chemistry of novel xanthophyll carotenoids. Am. J. Cardiol. 101, 95–100 (2008).
57. Faulk, R. M. & Southon, S. Challenges to understanding and measuring carotenoid bioavailability. Biochim. Biophys. Acta 1740, 95–100 (2005).
58. Itoh K, Mimura J & Yamamoto M. Discovery of the negative regulator of Nrf2, Keap1: An historical overview. Antioxid. Redox Signal 13, 1665–1678 (20109).
59. Bellezza I, Giambanco I, Minelli A, 0 & Donato R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta 1865, 721–733 (2018).
60. Stefanson, A. L. & Bakovic, M. Dietary regulation of Keap1/Nrf2/ARE pathway: focus on plant-derived compounds and trace minerals. Nutirents 6, 3777–3801 (2014).
61. Ben-Dor, A. et al. Carotenoids activate the antioxidant response element transcription system. Mol. Cancer Ther. 2005, 177–186 (2005).
62. Linnewiel K et al. Structure activity relationship of carotenoid derivatives in activation of the electrophile-antioxidant response element transcription system. Free Rad. Biol. Med. 47, 659–667 (2009).
63. Desai, K. & Sivakami, S. Purification and biochemical characterization of a superoxide dismutase from the soluble fraction of the cyanobacteium Spirulina platensis. World J. Micro. Biotech. 23, 1661–1666 (2007).
64. El Baky, H. H., El-Baz, F. K. & Baroty, G. E. Production of phenolic compounds from Spirulina maxima microalgae and its protective effects. Afr. J. Biotech. 8, 7059–7067 (2009).
65. Kong, B.-W. et al. RNA sequencing for global gene expression associated with muscle growth in a single male modern broiler line compared to a foundational Barred Plymouth Rock chicken line. BMC Genomics 18, (2017).
66. Khatri B et al. MicroRNA profiling associated with muscle growth in modern broilers compared to an unselected chicken breed. BMC Genomics 19, 683 (2018).
67. Palozza, P. Prooxidant actions of carotenoids in biologic systems. Nutr. Rev. 56, 257–265 (1998).
68. Palozza P et al. Beta-carotene at high concentrations induces apoptosis by enhancing oxy-radical production in human adenocarcinoma cells. Free Rad. Biol. Med. 30, 1000–1007 (2001).
69. Paolini M et al. Induction of cytochrome P450 enzymes and over-generation of oxygen radicals in beta-carotene supplemented rats. Carcinogenesis 22, 1483–1495 (2001).
70. Amengual J et al. A mitochondrial enzyme degrades carotenoids and protects against oxidative stress. FASEB J. 25, 948–959 (2011).
71. Johnson JD & Hill GE. Is carotenoid ornamentation linked to the inner mitochondria membrane potential? A hypothesis for the maintenance of signal honesty. Biochimie 21, 436–444 (2013).
72. Mayne, S. T. & Parker, R. S. Subcellular distribution of dietary beta-carotene in chick liver. Lipids 21, 164–169 (1986).
73. Barja, G. Mitochondrial free radical production and aging in mammals and birds. Ann. N.Y. Acad. Sci. 854, 224–238 (1998).
74. Karadas F, Erdogan S, Kor D, Oto G & Uluman M. The effects of different types of antioxidants (Se, Vitamin E and Carotenoids) in broiler diets on the growth performance, skin pigmentation and liver and plasma antioxidant concentrations. . Rev. Bras. Cienc. Avic. 18, 101–115 (2016).
75. Moreno J. A. et al. The distribution of carotenoids in hens fed on biofortified maize is influenced by feed composition, absorption, resource allocation and storage. Sci. Rep. 6, 35346 (2016).